Glucolípidos: clasificación, estructura y funciones
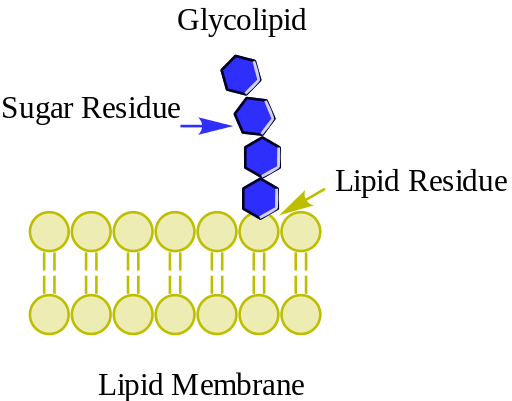
Los glucolípidos son lípidos de membrana con carbohidratos en sus grupos de cabeza polar. Presentan la distribución más asimétrica entre los lípidos de membrana, ya que se encuentran exclusivamente en la monocapa externa de las membranas celulares, siendo particularmente abundantes en la membrana plasmática.
Como la mayor parte de los lípidos de membrana, los glucolípidos poseen una región hidrofóbica compuesta por colas hidrocarbonadas apolares, y una cabeza o región polar, que puede estar conformada por diversas clases de moléculas, dependiendo del glucolípido en cuestión.
Los glucolípidos pueden encontrarse en organismos unicelulares como las bacterias y levaduras, así como en organismos tan complejos como animales y plantas.
En las células animales los glucolípidos están predominantemente compuestos por un esqueleto de esfingosina, mientras que en plantas los dos más comunes corresponden a diglicéridos y derivados de ácido sulfónico. En las bacterias también existen glicosil glicéridos y derivados de azúcares acilados.
En las plantas los glucolípidos están concentrados en las membranas cloroplasticas, mientras que en los animales abundan en la membrana plasmática. Junto con glicoproteínas y proteoglicanos, los glucolípidos forman parte importante del glucocálix, que es crucial para muchos procesos celulares.
Los glucolípidos, especialmente aquellos de las células animales, tienden a asociarse entre sí a través de enlaces de hidrógeno entre sus porciones carbohidratadas, y por fuerzas de van der Waals entre sus cadenas de ácidos grasos. Estos lípidos están presentes en estructuras membranales conocidas como balsas lipídicas, que poseen múltiples funciones.
Las funciones de los glucolípidos son varias, pero en eucariotas su ubicación en la cara externa de la membrana plasmática es relevante desde múltiples puntos de vista, especialmente en procesos de comunicación, adhesión y diferenciación celular.
Índice del artículo
Los glucolípidos son glicoconjugados que forman un grupo muy heterogéneo de moléculas, cuya característica común es la presencia de residuos sacáridos unidos por enlaces glucosídicos a una mitad hidrofóbica, que puede ser acil-glicerol, ceramida o prenil fosfato.
Su clasificación se basa en el esqueleto molecular que es puente de unión entre la región hidrofóbica y polar. Así pues, dependiendo de la identidad de este grupo se tienen:
Estos glucolípidos, al igual que los glicerolípidos, tienen un esqueleto diacilglicerol o monoalquil-monoacilglicerol al que se unen los residuos azúcares por enlaces glucosídicos.
Los glicoglicerolípidos son relativamente uniformes en cuanto a su composición de carbohidratos, pudiendo hallarse en su estructura residuos de galactosa o glucosa, de donde se desprende su clasificación principal, a saber:
- Galacto glicerolípidos: poseen residuos de galactosa en su porción carbohidratada. La región hidrofóbica consiste en una molécula de diacilglicerol o alquil-acilglicerol.
- Gluco glicerolípidos: estos tienen residuos de glucosa en su cabeza polar y la región hidrofóbica se compone únicamente de alquil-acilglicerol.
- Sulfo glicerolípidos: pueden ser o galacto glicerolípidos o gluco glicerolípidos con carbonos unidos a grupos sulfato, que les dan la característica de “acídicos” y los diferencian de los glicoglicerolípidos neutrales (galacto- y gluco glicerolípidos).
Estos lípidos poseen como molécula “esqueleto” una porción de ceramida que puede tener diferentes moléculas de ácidos grasos unidas.
Son lípidos sumamente variables, no solo en cuanto a la composición de sus cadenas hidrofóbicas, sino respecto a los residuos carbohidratados en su cabeza polar. Son abundantes en numerosos tejidos de mamíferos.
Su clasificación se basa en el tipo de sustitución o en la identidad de la porción sacárida, más que en la región compuesta por las cadenas hidrofóbicas. De acuerdo con los tipos de sustitución, la clasificación de estos esfingolípidos es la siguiente:
Glucoesfingolípidos neutros: aquellos que contienen en la porción sacárida hexosas, N-acetil hexosaminas y metil pentosas.
Sulfatidas: son los glucoesfingolípidos que contienen ésteres sulfato. Poseen carga negativa y son especialmente abundantes en las vainas de mielina en las células cerebrales. Los más comunes poseen un residuo de galactosa.
Gangliósidos: también conocidos como sialosil glucolípidos, son aquellos que contienen ácido siálico, por lo que también se conocen como glucoesfingolípidos acídicos.
Fosfoinositido-glucolípidos: el esqueleto está conformado por fosfoinositido-ceramidas.
Son lípidos usualmente reconocidos como anclas estables para proteínas en la bicapa lipídica. Son añadidos postraduccionalmente al extremo C-terminal de muchas proteínas que típicamente se encuentran mirando hacia la cara externa de la membrana citoplasmática.
Están compuestos por un centro glucano, una cola fosfolipídica y una porción fosfoetanolamina que los une.
Los glucolípidos pueden tener las porciones sacáridas unidas a la molécula por enlaces N- u O-glucosídicos, e incluso a través de enlaces no-glucosídicos, como los enlaces éster o amida.
La porción sacárida es sumamente variable, no solo en estructura sino en composición. Esta porción sacárida puede estar compuesta por mono-, di-, oligo- o polisacáridos de distintos tipos. Pueden tener aminoazúcares e incluso azúcares acídicos, simples o ramificados.
A continuación, una breve descripción de la estructura general de las tres principales clases de glucolípidos:
Como se mencionó con anterioridad, en los animales los glicoglicerolípidos pueden tener residuos de galactosa o de glucosa, fosfatados o no. Las cadenas de ácidos grasos en estos lípidos son de entre 16 y 20 átomos de carbono.
En los galacto glicerolípidos, la unión entre el azúcar y el esqueleto lipídico ocurre por enlaces β-glucosídicos entre el C-1 de la galactosa y el C-3 del glicerol. Los otros dos carbonos del glicerol están o bien esterificados con ácidos grasos o el C1 es sustituido por un grupo alquilo y el C2 con un grupo acilo.
Por lo general se observa un solo residuo de galactosa, aunque se ha reportado la existencia de digalactoglicerolípidos. Cuando se trata de un slufogalactoglicerolípido, normalmente el grupo sulfato se encuentra en el C-3 del residuo de galactosa.
La estructura de los gluco glicerolípidos es un poco diferente, sobre todo respecto al número de residuos de glucosa que puede ser de hasta 8 residuos unidos entre sí por enlaces de tipo α (1-6). La molécula de glucosa que sirve de puente con el esqueleto lipídico se une a este por un enlace α (1-3).
En los sulfoglucoglicerolípidos el grupo sulfato se une al carbono en posición 6 del residuo de glucosa terminal.
Al igual que los demás esfingolípidos, los glucoesfingolípidos derivan de una L-serina condensada con un ácido graso de cadena larga que forma una base esfingoide conocida como esfingosina. Cuando al carbono 2 de la esfingosina se une otro ácido graso se produce una ceramida, que es la base común para todos los esfingolípidos.
Dependiendo del tipo de esfingolípido estos están compuestos por residuos de D-glucosa, D-galactosa, N-acetil-D-galactosamina y N-acetilglucosamina, así como ácido siálico. Los gangliósidos son quizá los más diversos y complejos en cuanto a las ramificaciones de las cadenas oligosacáridas.
En estos glucolípidos los residuos del centro glucano (glucosamina y manosa) pueden ser modificados de diferentes maneras a través de la adición de grupos de fosfoetanolamina y otros azúcares. Esta variedad les proporciona una gran complejidad estructural que es importante para su inserción en la membrana.
Los cloroplastos de muchas algas y plantas superiores están enriquecidos de galacto glicerolípidos neutros que tienen propiedades similares a la de los cerebrósidos en los animales. Los mono- y digalactolípidos están β-unidos a una porción diglicérida, mientras que los sulfolípidos son derivados solo de α-glucosa.
En las bacterias, los glicosil glicéridos son estructuralmente análogos a los fosfoglicéridos animales, pero contienen residuos de carbohidratos unidos por glucosilación en la posición 3 del sn-1,2-diglicérido. Los derivados de azúcares acilados no contienen glicerol sino ácidos grasos unidos directamente a los azúcares.
Los residuos sacáridos más comunes entre los glucolípidos bacterianos son galactosa, glucosa y manosa.
En los animales, los glucolípidos tienen una importante función en la comunicación celular, diferenciación y proliferación, oncogénesis, repulsión eléctrica (en el caso de los glucolípidos polares), adhesión celular, entre otros.
Su presencia en muchas de las membranas celulares de animales, plantas y microorganismos da cuenta de su importante función, que está relacionada particularmente con las propiedades de las balsas lipídicas multifuncionales.
La porción carbohidratada de los glucoesfingolípidos es un determinante de la antigenicidad e inmunogenicidad de las células que lo portan. Puede estar implicada en procesos de reconocimiento intercelular, así como en actividades “sociales” celulares.
Los galacto glicerolípidos en las plantas, dada su abundancia relativa en las membranas vegetales, tienen una importante función en el establecimiento de las características de la membrana como estabilidad y actividad funcional de muchas proteínas de membrana.
La función de los glucolípidos en las bacterias también es diversa. Algunos de los glicoglicerolípidos son requeridos para mejorar la estabilidad de la bicapa. Sirven también como precursores de otros componentes de membrana y también soportan el crecimiento en anoxia o deficiencia de fosfato.
Las anclas GPI o los glucosidilfosfatidilinositoles también están presentes en las balsas lipídicas, participan de la transducción de señales, en la patogénesis de muchos microorganismos parasíticos y en la orientación de la membrana apical.
Se puede decir entonces que las funciones generales de los glucolípidos, tanto en plantas, como en animales y en bacterias, corresponden con el establecimiento de la estabilidad y fluidez de la membrana; participación en interacciones específicas lípido-proteína y reconocimiento celular.
1. Abdel-mawgoud, A. M., & Stephanopoulos, G. (2017). Simple glycolipids of microbes: Chemistry, biological activity and metabolic engineering. Synthetic and Systems Biotechnology, 1–17.
2. Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2015). Molecular Biology of the Cell (6th ed.). New York: Garland Science.
3. Ando, T., Imamura, A., Ishida, H., & Kiso, M. (2007). Synthesis of Glycolipids. Carbohydrate Research, 797–813.
4. Benson, A. (1964). Plant membrane lipids. Annu. Rev. Plant. Physiol., 15, 1–16.
5. Bronislaw, L., Liau, Y. U. N. H., & Slomiany, A. (1987). Animal glycoglycerolipids. Prog. Lipid Res., 26, 29–51.
6. Holzl, G., & Dormann, P. (2007). Structure and function of glycoglycerolipids in plants and bacteria. Prog. Lipid Res., 46, 225–243.
7. Honke, K. (2013). Biosynthesis and biological function of sulfoglycolipids. Proc. Jpn. Acad. Ser. B, 89(4), 129–138.
8. Kanfer, J., & Hakomori, S. (1983). Sphingolipid Biochemistry. (D. Hanahan, Ed.), Handbook of Lipid Research 3 (1st ed.).
9. Koynova, R., & Caffrey, M. (1994). Phases and phase transitions of the glycoglycerolipids. Chemistry and Physics of Lipids, 69, 181–207.
10. Law, J. (1960). Glycolipids. Annual Reviews, 29, 131–150.
11. Paulick, M. G., & Bertozzi, C. R. (2008). The Glycosylphosphatidylinositol Anchor: A Complex Membrane-Anchoring. Biochemistry, 47, 6991–7000.