Cloroplastos: características, funciones y estructura
Los cloroplastos son un tipo de orgánulos celulares delimitados por un sistema complejo de membranas, característicos de las plantas y las algas. En este plastidio se encuentra la clorofila, pigmento responsable de los procesos de fotosíntesis, del color verde de los vegetales y de permitir la vida autótrofa de estos linajes.
Además, los cloroplastos están relacionados con la generación de energía metabólica (ATP – adenosin trifosfato), síntesis de aminoácidos, vitaminas, ácidos grasos, componentes lípidos de sus membranas y reducción de nitritos. También posee un papel en la producción de sustancias de defensa contra patógenos.
Esta orgánulo fotosintético posee su propio genoma circular (ADN) y se plantea que, al igual que las mitocondrias, se originaron de un proceso de simbiosis entre un huésped y una bacteria fotosintética ancestral.
Índice del artículo
- 1 Origen
- 2 Características generales de los cloroplastos
- 3 Estructura (partes)
- 4 Funciones
- 5 Otros plastidios
- 6 Referencias
Los cloroplastos son organelas que poseen características de grupos de organismos muy distantes: las algas, las plantas y los procariotas. Esta evidencia sugiere que la organela se originó a partir de un organismo procariota con la capacidad de realizar fotosíntesis.
Se estima que el primer organismo eucariota, con la capacidad de realizar fotosíntesis, se originó hace unos 1.000 millones de años. Las pruebas indican que este importante salto evolutivo fue causado por la adquisición de una cianobacteria por parte de un huésped eucariota. Este proceso dio origen a distintos linajes de algas rojas, verdes y a las plantas.
Del mismo modo, se plantean eventos de simbiosis secundarias y terciarias en la que un linaje de eucariotas establece una relación simbiótica con otro eucariota fotosintético de vida libre.
Durante el curso de la evolución, el genoma de la supuesta bacteria se ha reducido y algunos de sus genes se han transferido e integrado al genoma del núcleo.
La organización del genoma de los cloroplastos actuales recuerda al de un procariota, sin embargo también posee atributos del material genético propio de eucariotas.
La teoría endosimbiótica fue propuesta por Lynn Margulis en una serie de libros publicados entre los años 60 y 80. No obstante, era una idea que ya venía manejándose desde los años 1900, propuesta por Mereschkowsky.
Esta teoría explica el origen de cloroplastos, mitocondrias y los cuerpos basales presentes en los flagelos. Según esta hipótesis, estas estructuras fueron alguna vez organismos procariotas libres.
No existe mucha evidencia que respalde el origen endosimbiótico de los cuerpos basales a partir de procariotas móviles.
En contraste, existe evidencia importante que apoya el origen endosimbiótico de mitocondrias a partir de α-Proteobacteria y de los cloroplastos a partir de cianobacterias. La evidencia más clara y contundente es la semejanza entre ambos genomas.
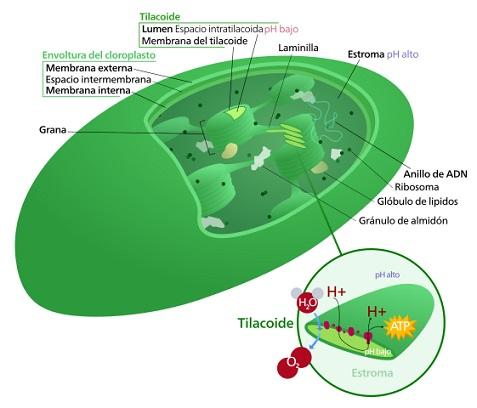
Los cloroplastos son el tipo de plastidios más conspicuos de las células vegetales. Son estructuras ovaladas rodeadas por membranas y en su interior ocurre el proceso más famoso de los eucariotas autótrofos: la fotosíntesis. Son estructuras dinámicas y poseen su propio material genético.
Generalmente se ubican en las hojas de las plantas. Una célula vegetal típica puede poseer de 10 a 100 cloroplastos, aunque el número es bastante variable.
Al igual que la mitocondria, la herencia de los cloroplastos de padres a hijos ocurre por parte de uno de los progenitores y no de ambos. De hecho, estos organelos son bastante similares a las mitocondrias en diversos aspectos, aunque más complejos.
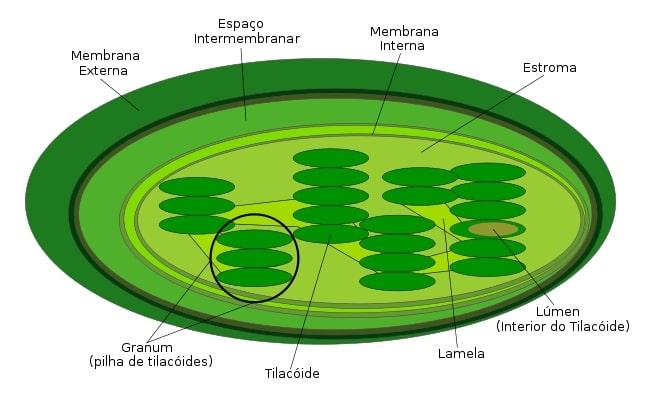
Los cloroplastos son organelas grandes, de 5 a 10 µm de longitud. Las características de esta estructura pueden ser visualizadas bajo un microscopio óptico tradicional.
Están rodeadas por una doble membrana lipídica. Además, poseen un tercer sistema de membranas internas, llamadas membranas del tilacoide.
Este último sistema membranoso forma un conjunto de estructuras similares a un disco, conocidas como tilacoides. La unión de tilacoides en pilas se denomina “grana” y están conectados entre sí.
Gracias a este triple sistema de membranas, la estructura interna del cloroplasto es compleja y se divide en tres espacios: el espacio intermembrana (entre las dos membranas externas), el estroma (encontrado en el cloroplasto y fuera de la membrana del tilacoide) y por último el lumen del tilacoide.
El sistema de membrana está relacionado con la generación de ATP. Al igual que las membranas de la mitocondria, es la membrana interna la que determina el paso de las moléculas al interior de la organela. La fosfaditilcolina y el fosfaditilglicerol son los lípidos más abundantes de las membranas del cloroplasto.
La membrana externa contiene una serie de poros. Por estos canales pueden ingresar libremente pequeñas moléculas. La membrana interna, por su parte, no permite el transito libre de este tipo de moléculas de bajo peso. Para que las moléculas ingresen, deben hacerlo por medio de transportadores específicos anclados a la membrana.
En algunos casos existe una estructura llamada retículo periférico, formada por una red de membranas, originada específicamente de la membrana interna del cloroplasto. Algunos autores las consideran únicas de plantas con metabolismo C4, aunque se han encontrado en plantas C3.
La función de estos túbulos y vesículas aún no está clara. Se propone que podrían contribuir al transporte rápido de metabolitos y proteínas dentro del cloroplasto o para incrementar la superficie de la membrana interna.
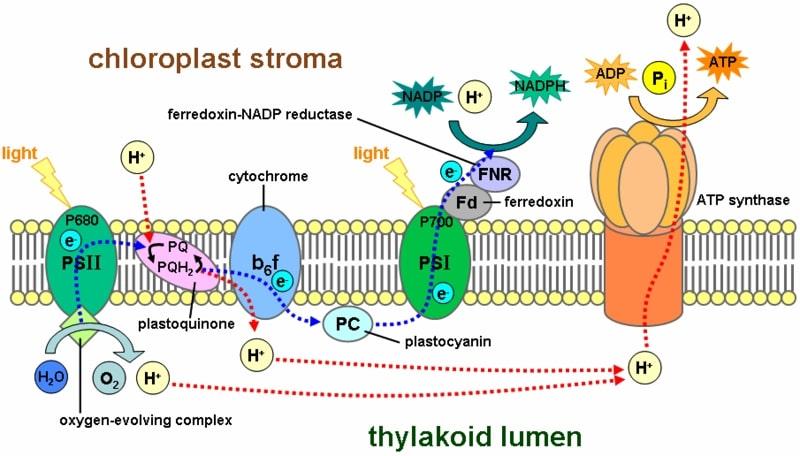
La cadena transportadora de electrones involucrada en los procesos fotosintéticos ocurre en este sistema de membranas. Los protones son bombeados a través de esta membrana, desde el estroma hacia el interior de los tilacoides.
Este gradiente resulta en la síntesis de ATP, cuando los protones son dirigidos nuevamente al estroma. Este proceso es equivalente al que ocurre en la membrana interna de la mitocondria.
La membrana del tilacoide está formada por cuatro tipos de lípidos: monogalactosil diacilglicerol, digalactosil diacilglicerol, sulfoquinovosil diacilglicerol y fosfatidilglicerol. Cada tipo cumple una función especial dentro de la bicapa lipídica de esta sección.
Los tilacoides son estructuras membranosas en forma de sacos o discos planos que se apilan en una “grana” (el plural de esta estructura es granum). Estos discos poseen un diámetro de 300 a 600 nm. En espacio interno del tilacoide se denomina lumen.
La arquitectura del apilamiento de tilacoides aún es debatida. Se proponen dos modelos: el primero es el modelo helicoidal, en la que los tilacoides se enrollan entre las granas en forma de hélice.
En contraste, el otro modelo propone una bifurcación. Esta hipótesis sugiere que las grana se encuentran formadas por bifurcaciones del estroma.
El estroma es el líquido gelatinoso que rodea a los tilacoides y se encuentra en la región interna del cloroplasto. Esta región corresponde al citosol de la supuesta bacteria que originó a este tipo de plastidio.
En esta zona se encuentra las moléculas de ADN y gran cantidad de proteínas y enzimas. Especificamente se encuentran las enzimas que participan en el ciclo de Calvin, para la fijación del anhídrido carbónico en el proceso fotosintético. También se pueden encontrar gránulos de almidón
En el estroma se encuentras los ribosomas de los cloroplastos, ya que estas estructuras sintetizan sus propias proteínas.
Una de las características más importantes de los cloroplastos es que poseen su propio sistema genético.
El material genético de los cloroplastos consiste en moléculas circulares de ADN. Cada organela posee múltiples copias de esta molécula circular de 12 a 16 kb (kilobases). Se organizan en estructuras llamadas nucleoides y consisten en 10 a 20 copias del genoma plastidico, junto con proteínas y moléculas de ARN.
El ADN del cloroplasto codifica para aproximadamente 120 a 130 genes. Estos resultan en proteínas y ARN relacionados con los procesos fotosintéticos como los componente del fotosistema I y II, la ATP sintasa y una de las subunidades de la Rubisco.
La Rubisco (ribulosa-1,5-bisfosfato carboxilasa/oxigenasa) es un complejo enzimático crucial en el ciclo de Calvin. De hecho, es considerada la proteína más abundante del planeta tierra.
Los ARN de transferencia y los ribosomales son usados en la traducción de ARN mensajesos que están codificados en el genoma del cloroplasto. Incluye los ARN ribosomales 23S, 16S, 5S y 4,5S y 30 ARN de transferencia. También codifica para 20 proteínas ribosomales y ciertas subunidades de la ARN polimerasa.
No obstante, ciertos elementos necesarios para el funcionamiento del cloroplasto están codificados en el genoma nuclear de la célula vegetal.
Los cloroplastos pueden ser considerados como importantes centro metabólicos en las plantas, donde ocurren múltiples reacciones bioquímicas gracias al amplio espectro de enzimas y proteínas ancladas a membranas que estas organelas contienen.
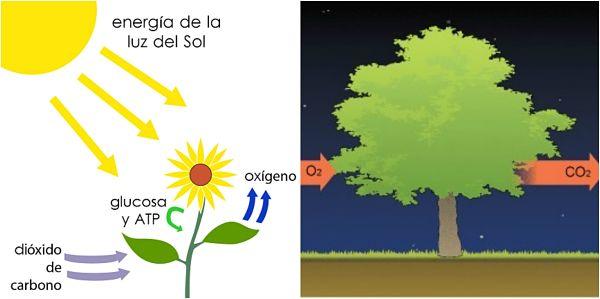
Tienen una función crítica en los organismos vegetales: es el lugar donde ocurren los procesos fotosintéticos, donde la luz solar es transformada en carbohidratos, teniendo como producto secundario el oxígeno.
En los cloroplastos también ocurre una serie de funciones secundarias de biosíntesis. A continuación discutiremos en detalle cada función:
La fotosíntesis ocurre gracias a la clorofila. Este pigmento se encuentra dentro de los cloroplastos, en las membranas de los tilacoides.
Está compuesto por dos partes: un anillo y una cola. El anillo contiene magnesio y es el responsable de la absorción de la luz. Puede absorber luz azul y luz roja, reflejando la zona verde del espectro de luz.
Las reacciones fotosintéticas ocurren gracias a la transferencia de electrones. La energía proveniente de la luz confiere energía al pigmento clorofila (se dice que la molécula es “excitada por la luz”), provocando un movimiento de estas partículas en la membrana de los tilacoides. La clorofila obtiene sus electrones de una molécula de agua.
Este proceso trae como consecuencia la formación de un gradiente electroquímico que permite la síntesis de ATP en el estroma. Esta fase también se conoce como “luminosa”.
La segunda parte de la fotosíntesis (o fase oscura) ocurre en el estroma y sigue en el citosol. Se conoce también como reacciones de fijación de carbono. En esta etapa, los productos de las reacciones anteriores son usados para construir carbohidratos a partir de CO2.
Además, los cloroplastos poseen otras funciones especializadas que permiten el desarrollo y el crecimiento de la planta.
En esta organela ocurre la asimilación de nitratos y sulfatos, y poseen las enzimas necesarias para la síntesis de aminoácidos, fitohormonas, vitaminas, ácidos grasos, clorofila y carotenoides.
Ciertos estudios han identificado un número importante de aminoácidos sintetizados por esta organela. Kirk y colaboradores estudiaron la producción de aminoácidos en los cloroplastos de Vicia faba L.
Estos autores encontraron que los aminoácidos sintetizados más abundantes fueron el glutamato, el aspartato y la treonina. Otro tipos, como alanina, serina y glicina también fueron sintetizados pero en menor cantidad. También fueron detectados los trece aminoácidos restantes.
Se han logrado aislar distintos genes que intervienen en la síntesis de lípidos. Los cloroplastos poseen las vías necesarias para la síntesis de lípidos isoprenoides, esencial para la producción de clorofila y otros pigmentos.
Las plantas no cuentan con un sistema inmune desarrollado similar al de los animales. Por ello, las estructuras celulares deben producir sustancias antimicrobianas para poder defenderse contra agentes perjudiciales. Para este propósito, las plantas pueden sintetizar especies reactivas del oxígeno (ROS) o ácido salicílico.
Los cloroplastos están relacionados con la producción de estas sustancias que eliminan posibles agentes patógenos que ingresan a la planta.
Igualmente, funcionan como un “sensores moleculares” y participan en mecanismos de alerta, comunicando la información a otras organelas.
Los cloroplastos pertenecen a una familia de organelas vegetales llamadas plastidios o plastos. Los cloroplastos se diferencian principalmente del resto de los plastidios por poseer el pigmento clorofila. Los demás plastidios son:
-Los cromoplastos: estas estructuras contienen carotenoides, están presentes en flores y flores. Gracias a estos pigmentos, las estructuras vegetales poseen colores amarillos, naranjas y rojos.
-Los leucoplastos: estos plastidios no contienen pigmentos y por lo tanto son blancos. Sirven como reserva y se encuentran en órganos que no reciben luz directa.
-Los amiloplastos: contienen almidón y se encuentran en raíces y tubérculos.
Los plastidios se originan de estructuras llamadas protoplastidios. Una de las características más sorprendentes de los plastidios es su propiedad para cambiar de tipo, aunque ya estén en etapa madura. Este cambio se desencadena por señales ambientales o intrínsecas de la planta.
Por ejemplo, los cloroplastos son capaces de dar origen a los cromoplastos. Para este cambio, la membrana del tilacoide se desintegra y los carotenoides son sintetizados.
- Allen, J. F. (2003). Why Chloroplasts and Mitochondria Contain Genomes. Comparative and Functional Genomics, 4(1), 31–36.
- Cooper, G. M (2000). The cell: Molecular approach. Second edition. Sinauer Associates
- Daniell, H., Lin, C.-S., Yu, M., & Chang, W.-J. (2016). Chloroplast genomes: diversity, evolution, and applications in genetic engineering. Genome Biology, 17, 134.
- Gracen, V. E., Hilliard, J. H., Brown, R. H., & West, S. H. (1972). Peripheral reticulum in chloroplasts of plants differing in CO 2 fixation pathways and photorespiration. Planta, 107(3), 189-204.
- Gray, M. W. (2017). Lynn Margulis and the endosymbiont hypothesis: 50 years later. Molecular Biology of the Cell, 28(10), 1285–1287.
- Jensen, P. E., & Leister, D. (2014). Chloroplast evolution, structure and functions. F1000Prime Reports, 6, 40.
- Kirk, P. R., & Leech, R. M. (1972). Amino Acid Biosynthesis by Isolated Chloroplasts during Photosynthesis . Plant Physiology, 50(2), 228–234.
- Kobayashi, K., & Wada, H. (2016). Role of lipids in chloroplast biogenesis. In Lipids in Plant and Algae Development (pp. 103-125). Springer, Cham.
- Sowden, R. G., Watson, S. J., & Jarvis, P. (2017). The role of chloroplasts in plant pathology. Essays in biochemistry, EBC20170020.
- Wise, R. R., & Hoober, J. K. (2007). The structure and function of plastids. Springer Science & Business Media.