Nastias: tipos, características y ejemplos
Las nastias, nastismos o movimientos násticos son una forma de movimiento de las plantas que resulta de la percepción de un estímulo externo en una dirección, pero donde la dirección del movimiento resultante es independiente de la del estímulo percibido. Ocurren prácticamente en todos los órganos de las plantas: hojas, tallos y ramas, flores, zarcillos y raíces.
Entre los mecanismos que tienen las plantas para adaptarse al medio que las rodea se encuentran algunas formas de movimientos que, reversible o irreversiblemente, resultan de la percepción de estímulos lumínicos, térmicos, químicos, hídricos, táctiles, gravitacionales, producto de heridas causadas por herbívoros al alimentarse, entre otros.

El movimiento en las plantas está clasificado tradicionalmente en dos tipos: los tropismos y las nastias. Los tropismos, a diferencia de las nastias, se definen funcionalmente como movimientos o respuestas de crecimiento de órganos vegetales frente a estímulos físicos y están relacionados directamente con la dirección en la que estos se perciben.
Tanto nastias como tropismos pueden ser el resultado de movimientos por crecimiento o por cambios de turgencia en las células del órgano que se mueve, por lo que algunos movimientos podrían considerarse reversibles y otros irreversibles, según sea el caso.
Charles Darwin en su obra del año 1881 –El poder del movimiento en las plantas– describió los movimientos vegetales producto de cambios medioambientales, especialmente aquellos relacionados con respuestas trópicas. No obstante, los mecanismos subyacentes a estos movimientos han sido descritos por diversos autores desde entonces hasta el tiempo presente.
Índice del artículo
Una planta puede recibir gran diversidad de estímulos para los cuales puede desencadenar gran variedad de respuestas. La clasificación de los distintos movimientos násticos se ha hecho principalmente en base a la naturaleza de los estímulos, sin embargo, la descripción científica de los mecanismos de respuesta presenta muchas ambigüedades.
Entre los tipos de nastias más conocidas se encuentran:
- Nictinastia: cuando las hojas de algunas especies de plantas leguminosas se expanden completamente durante el día y se pliegan o cierran durante la noche.
- Tigmonastia/Seismonastia: movimientos que resultan de estímulos por contacto físico directo en ciertos órganos de algunas especies.
- Termonastia: movimientos reversibles dependientes de fluctuaciones térmicas.
- Fotonastia: se considera un tipo especial de fototropismo; las hojas de algunas especies en condiciones de alta intensidad lumínica pueden disponerse paralelamente a la incidencia de luz.
- Epinastia e hiponastia: son movimientos foliares que poseen algunas especies frente a condiciones de extrema humedad en las raíces o altas concentraciones de sal en el suelo. La epinastia tiene que ver con crecimiento exagerado de la región adaxial mientras que la hiponastia se refiere al crecimiento de la región abaxial de la lámina foliar.
- Hidronastia: movimiento de ciertos órganos vegetales que depende de estímulos hídricos.
- Quimionastia: respuesta de movimiento relacionada con gradientes de concentración de algunas sustancias químicas. Algunos autores hacen referencia más bien a movimientos internos y vías de señalización.
- Gravinastia/Geonastia: movimiento temporal reversible de algunas plantas en respuesta a los estímulos gravitacionales.
Muchos de los movimientos násticos dependen de la existencia de un órgano particular: el pulvínulo. Los pulvínulos son órganos motores especializados ubicados en la base de los pecíolos de las hojas simples, y pecíolos y foliolos en hojas compuestas.
Anatómicamente hablando, se componen de un cilindro central, rodeado por capas de colénquima, y una zona cortical motora que posee células parenquimáticas susceptibles a cambios de tamaño y forma.
Las células de la corteza pulvinular que cambian de tamaño y forma se conocen como células motoras, entre las que se distinguen las células motoras extensoras y las flexoras. Normalmente el movimiento de estas depende de cambios de turgencia debidos a la entrada y/o salida del agua del protoplasto.
A continuación se presenta una breve descripción de las nastias cuyos casos podrían considerarse clásicos ejemplos.
Fueron descubiertos inicialmente en Mimosa púdica y son muy comunes en leguminosas. Tienen que ver con el movimiento “rítmico” de las hojas, que se cierran por las noches y se expanden completamente durante el día. Los más estudiados han sido los de Albizzia julibrissim, A. lophantha, Samanea saman, Robinia pseudoacacia y Phaseolus coccineus.
El fenómeno es muy conocido en las plantas y se piensa que tiene razones adaptativas: la expansión de las láminas foliares durante el día permite captar un máximo de energía lumínica durante la exposición al sol, mientras que el cierre por la noche busca evitar una pérdida calórica importante.
Cuando las hojas están expandidas los pulvínulos están en posición horizontal (diurna) y cuando están cerradas tienen forma de “U” (nocturna), o que se relaciona con un aumento de turgencia en las células extensoras durante la apertura, y con un incremento de turgencia en las células flexoras durante el cierre.
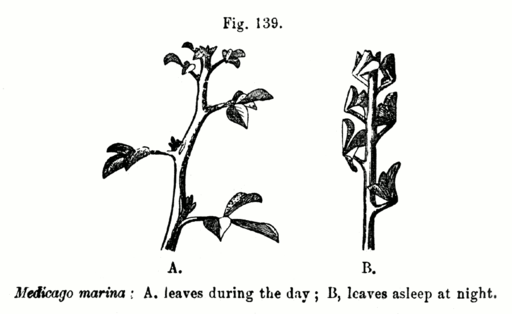
Tales cambios de turgencia se dan por el movimiento de agua que depende del movimiento intracelular de iones como el K+ y el Cl-, el malato y otros aniones.
El K+ ingresa a las células motoras por un aumento en la carga negativa en la cara interna de la membrana citoplasmática, que se consigue por medio de la acción de ATPasas encargadas de expulsar protones del citoplasma.
La pérdida de turgencia se da por la inactivación de la bomba de protones, lo que despolariza la membrana y activa los canales de potasio, promoviendo la salida de este ion hacia el apoplasto.
Estos movimientos dependen de la acción de fotorreceptores compuestos por fitocromos, en vista de que experimentos han demostrado que una radiación prolongada estimula la apertura foliar.
El movimiento nictinástico posee cierta “ritmicidad”, pues plantas sometidas a oscuridad permanente presentan estos movimientos cada 24h, por lo que una especie de “reloj biológico” debe participar en la regulación de los cambios de turgencia en las células motoras de los pulvínulos.
Una de las respuestas tigmonásticas más populares en la literatura es la que presenta la planta carnívora Dionaea muscipula o “Atrapamoscas de Venus”, donde los insectos quedan atrapados en sus hojas bilobuladas abisagradas.
Cuando un insecto trepa hacia la superficie ventral de la hoja y se topa con tres delicados pelos que disparan la respuesta motora, se generan señales eléctricas intercelulares e inicia el alargamiento diferencial de las células de cada lóbulo foliar, lo que resulta en el cierre de la “trampa” en menos de un segundo.

La carnivoría le da a D. muscipula suficiente nitrógeno para subsistir, por lo que estas plantas pueden asentarse sin problema en suelos pobres en este mineral. Es importante resaltar que este movimiento es muy específico, lo que quiere decir que estímulos como las gotas de lluvia o fuertes vientos no disparan el cierre de los lóbulos.
Otra planta carnívora, Drosera rotundifolia, posee cientos de tentáculos mucilaginosos en la superficie de sus hojas modificadas, lo que llama la atención de cientos de presas potenciales, que quedan atrapadas en el mucílago de los “tentáculos”.
Los tentáculos sensoriales detectan la presencia de la presa y los tentáculos adyacentes se inclinan hacia el que ha sido estimulado, formando una trampa en forma de copa que atrapa al insecto en su interior.
Se piensa que ocurre un crecimiento diferencial que está controlado por cambios en los niveles de auxina ya que la adición de auxinas exógenas dispara el cierre de las hojas, y al añadir bloqueadores del transporte de la misma hormona, se inhibe el movimiento.
Mimosa púdica también es protagonista de los movimientos tigmonásticos mejor descritos. El toque de uno de sus foliolos promueve el cierre inmediato de sus hojas compuestas.
Se piensa que esta respuesta a estímulos táctiles puede servir para ahuyentar posibles predadores o como mecanismo de defensa que permite la exposición de sus espinas defensivas.
El plegamiento de las hojas depende de cambios de turgencia. Los pulvínulos en este caso pierden turgor, concretamente, las células flexoras se estiran como respuesta de la pérdida de volumen de las células extensoras.

El cambio de volumen ocurre por una descarga de sacarosa en el floema, que obliga el transporte osmótico de agua y el transporte pasivo de iones potasio y cloro.
En este movimiento también se genera un gradiente electroquímico gracias a la participación de bombas de protones en la membrana (ATPasas). Están involucrados factores de crecimiento, citoesqueleto y filamentos de actina, entre otros.
Se ha detallado en flores de Crocus y en tulipanes. Ocurre por un crecimiento diferencial en los lados opuestos de los pétalos que reaccionan al estímulo térmico y no por cambios de turgencia. La diferencia de respuesta se da puesto que los dos costados del órgano tienen óptimos de crecimiento a temperaturas muy diferentes.

Durante este movimiento no ocurren cambios significativos de los valores osmóticos, de pH o de permeabilidad de los protoplastos. También se han observado aumentos significativos de CO2 intracelular que parece ser el factor que sensibiliza los tejidos a los cambios de temperatura.

Este movimiento es independiente de la intensidad lumínica y estrictamente dependiente del incremento en la temperatura. La convención entre diferentes autores es que la variación térmica debe ser entre 0.2°C y 0.5°C para observar un movimiento de las flores. Una caída de la temperatura de la misma magnitud provoca su cierre.
- Azcón-Bieto, J., & Talón, M. (2008). Fundamentos de fisiología vegetal (2da ed.). Madrid: McGraw-Hill Interamericana de España.
- Braam, J. (2005). In touch: plant responses to mechanical stimuli. New Phytologist, 165, 373–389.
- Brauner, L. (1954). Tropisms and Nastic movements. Annu. Rev. Plant. Physiol., 5, 163–182.
- Brown, A. H., Chapman, D. K., & Liu, S. W. W. (1974). Comparison of Leaf Epinasty Induced by Weightlessness or by Clinostat Rotation. Bioscience, 24(9), 518–520.
- Dubetz, S. (1969). An unusual photonastism induced by drought in Phaseolus vulgaris. Canadian Journal of Botany, 47, 1640–1641.
- Dumais, J., & Forterre, Y. (2012). “Vegetable Dynamics”: The Role of Water in Plant Movements. Annu. Rev. Fluid Mech., 44, 453–478.
- Enright, J. T. (1982). Sleep movements of leaves: In defense of Darwin’s interpretation. Oecologia, 54(2), 253–259.
- Esmon, C. A., Pedmale, U. V, & Liscum, E. (2005). Plant tropisms : providing the power of movement to a sessile organism. Int. J. Dev. Biol., 49, 665–674.
- Firn, R. D., & Myers, A. B. (1989). Plant movements caused by differential growth- unity of diversity of mechanisms? Environmental and Experimental Botany, 29, 47–55.
- Guo, Q., Dai, E., Han, X., Xie, S., Chao, E., & Chen, Z. (2015). Fast nastic motion of plants and bioinspired structures. J. R. Soc. Interface, 12.
- Hayes, A. B., & Lippincott, J. A. (1976). Growth and Gravitational Response in the Development of Leaf Blade Hyponasty. American Journal of Botany, 63(4), 383–387.
- Koukkari, W. L., & Hillman, W. S. (1968). Pulvini as the Photoreceptors in the Phytochrome Effect on Nyctinasty in Albizzia julibrissin. Plant Physiology, 43(5), 698–704.
- Sandalio, L. M., Rodríguez-Serrano, M., & Romero-Puertas, M. C. (2016). Leaf epinasty and auxin: A biochemical and molecular overview. Plant Science. Elsevier Ireland Ltd.
- Schildknecht, H. (1983). Turgorins, Hormones of the Endogenous Daily Rhythms of Higher Organized Plants—Detection, Isolation, Structure, Synthesis, and Activity. Angewandte Chemie International Edition in English, 22(9), 695–710.
- Ueda, M., Takada, N., & Yamamura, S. (2001). Molecular approach to the nyctinastic movement of the plant controlled by a biological clock. International Journal of Molecular Sciences, 2(4), 156–164.
- Wood, W. M. L. (1953). Thermonasty in Tulip and Crocus Flowers. Journal of Experimental Botany, 4(10), 65–77.