Renina: estructura, producción, secreción, funciones
La renina, también conocida como angiotensinogenasa, es una aspartil proteasa que tiene importantes implicaciones en la homeostasis de electrolitos y el control de la presión sanguínea en los mamíferos.
Esta proteína es secretada desde el riñón al torrente circulatorio y es responsable del aumento de la presión arterial en animales experimentales cuando se les inyecta extractos de riñón.
Como se trata de una sustancia producida por un tejido y secretada a la circulación con un blanco alejado de su sitio de producción, la renina es considerada como una hormona.
Las hormonas pueden ser proteínas o polipéptidos, tener origen esteroideo o derivar del aminoácido tirosina. La renina es una hormona de naturaleza proteica y su acción catalítica implica el clivaje enzimático de otras proteínas (es una proteasa).
Esta hormona fue descubierta a finales de la década de 1890. No obstante, no fue sino a finales de 1990 que se determinó con exactitud su origen fisiológico y su estructura molecular.
Índice del artículo
La renina humana es una glicoproteína con actividad enzimática y un peso molecular de poco más de 37 kDa. La molécula está formada por dos dominios separados por una hendidura profunda dentro de la cual se encuentra su sitio activo.
Ambos dominios de la renina son similares en secuencia y están compuestos principalmente por láminas β-plegadas.
Diferentes análisis de la secuencia de esta proteína revelan que tiene más de 30 residuos aminoacídicos básicos entre los que se incluyen varias argininas, lisinas e histidinas.
Además, se sabe que a lo largo de la estructura se encuentran centro hidrofóbicos y grandes superficies hidrofílicas que le proporcionan a la proteína estabilidad en diferentes contextos.
El sitio activo de la enzima se localiza en la hendidura formada por los dos dominios y los aminoácidos esenciales para la catálisis son dos residuos de ácido aspártico en las posiciones 38 y 226, razón por la cual esta es una “aspartil” proteasa.
La renina se produce en el aparato yuxtaglomerular del riñón, una estructura especializada que se encuentra en el sitio de contacto entre el túbulo contorneado distal y su glomérulo de origen.
Este aparato consta de tres componentes: las células granulares, las células mesangiales extraglomerulares y la mácula densa.
La mácula densa está formada por una hilera de células epiteliales cúbicas unidas de manera muy compacta que tapizan el tubo en el sitio de contacto con el glomérulo y que se considera el inicio del túbulo contorneado distal.
Las células mesangiales extraglomerulares se encuentran formando una región triangular entre la arteriola aferente, la arteriola eferente y la mácula densa, se consideran una extensión de las células mesangiales glomerulares. También se llaman células agranulares.
Las células granulares se denominan células yuxtaglomerulares y se ubican en las paredes de las arteriolas aferentes y eferentes y en la zona de las células mesangiales extraglomerulares.
Estas células granulares reciben dicha denominación por la presencia de gránulos de secreción en su citoplasma. Gránulos que contienen renina,así como un precursor de la renina, la pro-renina, que se forma a partir de la pre-pro-renina.
La pre-pro-renina es una prehormona que en el humano tiene 406 aminoácidos. Esta prehormona sufre un clivaje proteolítico post-traduccional, con lo que pierde una secuencia de 23 residuos en su extremo amino terminal.
El clivaje de la pre-pro-renina la convierte en pro-renina, de 383 aminoácidos. El clivaje posterior de otra secuencia en el extremo N-terminal de la pro-renina es lo que dirige la formación de la renina, una proteasa activa de 340 aminoácidos.
Tanto la pro-renina como la renina pueden ser secretadas a la circulación, pero muy poca pro-renina es convertida en renina activa en este tejido conectivo. Las enzimas encargadas de la conversión de la pro-renina en renina se conocen como calicreínas y catepsinas.
Una vez la renina es secretada a la circulación, tiene una vida media no mayor de 80 minutos y la secreción es altamente regulada.
Además del riñón, la renina puede ser producida por otros tejidos u órganos como los testículos, los ovarios, las paredes de las arteriolas, la corteza suprarrenal, la hipófisis, el cerebro, el líquido amniótico y otros.
Aunque es aplicable para muchos animales, estudios que implican la remoción de los riñones demuestran que la actividad renina en circulación cae drásticamente a niveles muy cercanos a cero.
La secreción de la renina incrementa por una serie de estímulos que aparecen cuando disminuye el volumen del líquido extracelular, cuando disminuye la presión arterial o cuando aumenta la actividad simpática en los nervios renales.
Se han descrito varios factores relacionados con la regulación de la secreción de renina:
– La presión de perfusión renal detectada por los barorreceptores (receptores de estiramiento) de la arteriola aferente
– Cambios en el volumen y en la composición del líquido que llega a la mácula densa
– Actividad de los nervios simpáticos renales
– Las prostaglandinas
– El péptido natriurético auricular.
El mecanismo barorreceptor de la arteriola aferente provoca una disminución de la secreción de renina cuando ocurre un aumento en la presión de la arteriola aferente a nivel del aparato yuxtaglomerular. Su secreción aumenta cuando disminuye la actividad barorreceptora al caer la presión.
Otro sensor relacionado con la regulación de la secreción de renina se encuentra en la mácula densa. Cuanto mayor sea la tasa de reabsorción de Na+ y Cl- y la concentración de estos electrolitos en el líquido que alcanza la mácula densa, menor será la secreción de renina y viceversa.
El incremento de la actividad de los nervios simpáticos renales, así como de las catecolaminas circulantes por medio de la norepinefrina liberada en las terminaciones simpáticas en las células yuxtaglomerulares, aumenta la secreción de renina.
Las prostaglandinas, específicamente las prostaciclinas, estimulan la secreción de renina por efecto directo sobre las células granulares del aparato yuxtaglomerular.
La angiotensina II, por un efecto de retroalimentación negativa, inhibe la secreción de renina por efecto directo sobre las células granulares. Otra hormona como la vasopresina inhibe la secreción de renina.
El péptido natriurético auricular (PNA), que se produce en el músculo auricular cardíaco, inhibe la secreción de renina.
El efecto sumado de todos los factores estimuladores e inhibidores es lo que determina la tasa de secreción de renina. La renina se segrega en la sangre renal y luego sale de los riñones para circular por todo el cuerpo. Sin embargo, una pequeña cantidad de renina se queda en los líquidos renales.
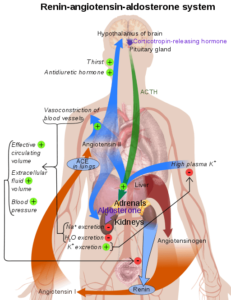
La renina es una enzima que por sí misma no posee funciones vasoactivas. La única función conocida de la renina es la de cortar el angiotensinógeno en el extremo amino terminal, generando un decapéptido denominado Angiotensina I.
El angiotensinógeno es una glicoproteína del grupo de las α2 globulinas sintetizada por el hígado y está presente en la sangre circulante.
Dado que la angiotensina I tiene una actividad vasopresora muy pobre y esta debe ser procesada “aguas abajo” por otra proteasa, la renina participa en los pasos iniciales de la regulación de la presión arterial, en un sistema que se conoce como renina-angiotensina.
La angiotensina II tiene una vida media muy corta (entre 1 y 2 minutos). Es metabolizada rápidamente por varias peptidasas que la van fragmentando y algunos de esos fragmentos, como la Angiotensina III conservan alguna actividad vasopresora.
Las funciones generales del sistema renina –angiotensina son múltiples y pueden resumirse como sigue:
– Constricción arteriolar y aumento en la presión sistólica y diastólica. La angiotensina II es de cuatro a ocho veces más potente que la noradrenalina para esta función.
– Incremento de la secreción de aldosterona por efecto directo de la Angiotensina II sobre la corteza suprarrenal. El sistema renina-angiotensina es el principal regulador de la secreción de aldosterona.
– Facilita la secreción de norepinefrina por efecto directo sobre las neuronas post-ganglionares simpáticas.
– Influye sobre contracción de las células mesangiales lo que disminuye la tasa de filtración glomerular y, por efecto directo sobre los túbulos renales, incrementa la reabsorción de sodio.
– A nivel cerebral, este sistema disminuye la sensibilidad del reflejo barorreceptor lo que potencia el efecto vasopresor de la Angiotensina II.
– La Angiotensina II estimula la ingesta de agua al promover los mecanismos de la sed. Aumenta la secreción de vasopresina y de hormona ACTH.
El sistema renina-angiotensina tiene, por tanto, un rol importante en las patologías hipertensivas, sobre todo aquellas de origen renal.
Es así como la constricción de una de las arterias renales genera una hipertensión sostenida que puede revertirse si se extrae el riñón isquémico (defectuoso) o se libera a tiempo la constricción arterial renal.
Un aumento en la producción de renina se asocia, generalmente, con la constricción unilateral de la arteria renal que conecta uno de los riñones, lo que ocasiona hipertensión. Esta condición clínica puede deberse a defectos congénitos u otras anormalidades de circulación renal.
La manipulación farmacológica de este sistema, además del uso de bloqueadores de los receptores de la angiotensina II, son las herramientas fundamentales para el tratamiento de la hipertensión arterial.
La hipertensión arterial es una enfermedad silente y progresiva que afecta a gran parte de la población mundial, especialmente a adultos mayores de 50 años.
- Akahane, K., Umeyama, H., Nakagawa, S., Moriguchi, I., Hirose, S., Iizuka, K., & Murakami, J. (1985). Three-dimensional Structure of Human Renin. Hypertension, 7(1), 3–12.
- Davis, J., & Freeman, R. (1976). Mechanisms Regulating Renin Release. Physiological Reviews, 56(1), 1–56.
- Guyton, A., & Hall, J. (2006). Textbook of Medical Physiology (11th ed.). Elsevier Inc.
- Hackenthal, E., Paul, M., Ganten, D., & Taugner, R. (1990). Morphology, Physiology , and Molecular Biology of Renin Secretion. Physiological Reviews, 70(4), 1067–1116.
- Morris, B. (1992). Molecular Biology of renin. I: Gene and protein structure, synthesis and processing. Journal of Hypertension, 10, 209–214.
- Murray, R., Bender, D., Botham, K., Kennelly, P., Rodwell, V., & Weil, P. (2009). Harper’s Illustrated Biochemistry (28th ed.). McGraw-Hill Medical.
- West, J. (1998). Bases Fisiológicas de la Práctica Médica (12ava ed.). México D.F.: Editorial Médica Panamericana.