Globósidos: estructura, biosíntesis, funciones y patologías
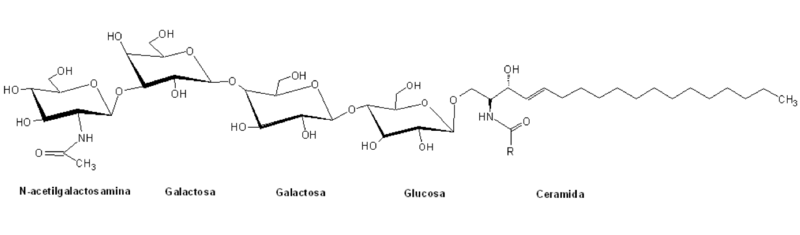
Los globósidos son un tipo de esfingolípidos pertenecientes a la heterogénea familia de los glucoesfingolípidos y están caracterizados por tener en sus estructuras un grupo polar compuesto por glicanos de compleja estructura unidos a un esqueleto de ceramida por un enlace B-glicosídico.
Se clasifican dentro de la serie “globo” de los glucoesfingolípidos por la presencia de una estructura central de la forma general Galα4Galβ4GlcβCer, y generalmente su nomenclatura se basa en el número y el tipo de residuos azucarados de las cabezas polares.
A diferencia de otros esfingolípidos, los globósidos son constituyentes normales de las membranas celulares de órganos sistémicos no nerviosos de muchos mamíferos. Por ejemplo los riñones, el intestino, los pulmones, las glándulas suprarrenales y los eritrocitos.
Como todos los lípidos de membrana, los globósidos tienen importantes funciones estructurales en la formación y ordenamiento de las bicapas lipídicas.
Sin embargo y a diferencia de sus contrapartes acídicas o fosforiladas, la función de los globósidos no se relaciona tanto CON la producción de moléculas señalizadoras, sino más bien con su participación como parte de los glicoconjugados en la membrana plasmática.
Índice del artículo
Comparten algunas similitudes estructurales y funcionales con los demás miembros del grupo de los glucoesfingolípidos: cerebrósidos, gangliósidos y sulfatidas; entre ellas la composición del esqueleto principal y los subproductos de su metabolismo.
Sin embargo, los globósidos difieren de los glucoesfingolípidos acídicos (como los gangliósidos) respecto a la carga de sus grupos polares carbohidratados, ya que son eléctricamente neutros a pH fisiológico, lo que parece tener fuertes implicaciones en sus funciones como parte de la matriz extracelular.
Estos grupos de cabeza polar tienen normalmente más de dos moléculas de azúcar, entre los que comúnmente se encuentran la D-glucosa, la D-galactosa y la N-acetil-D-galactosamina, y en menor proporción la fucosa y la N-acetilglucosamina.
Tal y como lo que ocurre con los demás esfingolípidos, los globósidos pueden ser moléculas muy diversas, bien sea tomando en cuenta las múltiples combinaciones de ácidos grasos unidos al esqueleto de esfingosina o las posibles variaciones de las cadenas de oligosacáridos de la porción hidrofílica.
La ruta comienza con la síntesis de ceramida en el retículo endoplasmático (ER). Primero se forma el esqueleto de esfingosina por condensación de una L-serina y un palmitoil-CoA.
La ceramida se genera posteriormente por acción de las enzimas ceramidas sintasas, que condensan otra molécula de ácido graso-CoA con el esqueleto de esfingosina en el carbono de la posición 2.
Aún en el ER, las ceramidas producidas pueden ser modificadas por la adición de un residuo de galactosa para formar galacto ceramidas (GalCer), o pueden en su lugar ser transportadas al complejo de Golgi bien por la acción de proteínas de transferencia de ceramidas (CERT) o por medio de transporte vesicular.
En el complejo de Golgi las ceramidas pueden ser glicosiladas para producir gluco ceramidas (GlcCer).
La GlcCer es producida en la cara citosólica del Golgi temprano. Luego puede ser transportada hacia la cara luminal del complejo y ser glicosilada subsecuentemente por enzimas glicosidasas específicas que generan glucoesfingolípidos más complejos.
Los precursores comunes de todos los glucoesfingolípidos son sintetizados en el complejo de Golgi por acción de las glicosiltransferasas a partir de GalCer o GlcCer.
Estas enzimas transfieren carbohidratos específicos a partir de los azúcares nucleótidos adecuados: UDP-glucosa, UDP-galactosa, CMP-ácido siálico, etc.
Cuando la GlcCer pasa por el sistema de tráfico vesicular del Golgi es galactosilada para producir lactosilceramida (LacCer). La LacCer es el punto de ramificación a partir del cual son sintetizados los precursores de los demás glucoesfingolípidos, es decir, la molécula a la cual se adicionan posteriormente más residuos azúcares polares neutros. Estas reacciones son catalizadas por globósido-sintasas específicas.
Estos lípidos se encuentran principalmente en tejidos humanos. Al igual que muchos glucoesfingolípidos, los globósidos están enriquecidos en la cara externa de la membrana plasmática de muchas células.
Son particularmente importantes en los eritrocitos humanos, donde representan el tipo principal de glucolípido de la superficie celular.
Además, como se ha señalado anteriormente, son parte del conjunto de glicoconjugados de las membranas plasmáticas de muchos órganos no-nerviosos, principalmente los riñones.
Las funciones de los globósidos no han sido esclarecidas completamente hasta la fecha, pero se sabe que algunas especies incrementan la proliferación y motilidad celular, en contraste con la inhibición de estos eventos causada por algunos gangliósidos.
Un globósido tetra glicosilado, el Gb4 (GalNAcβ3Galα4Galβ4GlcβCer), funciona en el reconocimiento sitio-sensible de perturbaciones estructurales de los eritrocitos durante los procesos de adhesión celular.
Estudios recientes han determinado la implicación del Gb4 en la activación de proteínas ERK en líneas celulares de carcinomas, lo que podría significar su participación en la iniciación tumoral. Estas proteínas pertenecen a la cascada de señalización de proteínas quinasas activadas por mitógenos (MAPK), que consisten en los elementos Raf, MEK y ERK.
Se ha reportado su participación como receptores para algunas toxinas bacterianas de la familia Shiga, concretamente el globósido Gb3 (Galα4Galβ4GlcβCer), también conocido como CD77, expresado en células B inmaduras; también como receptores para el factor de adhesión al VIH (gp120) y parecen tener implicaciones en ciertos tipos de cáncer y otras enfermedades.
Existen numerosos tipos de lipidosis en los seres humanos. Los globósidos y sus rutas metabólicas están relacionados con dos enfermedades en particular: la enfermedad de Fabry y la enfermedad de Sandhoff.
Se refiere a un desorden sistémico hereditario ligado al sexo, visto por primera vez en pacientes con múltiples manchas violetas en la región umbilical. Afecta órganos como los riñones, el corazón, los ojos, las extremidades, parte del sistema gastrointestinal y nervioso.
Es producto de un defecto metabólico en la enzima ceramida trihexosidasa, encargada de la hidrólisis de la trihexosiceramida, un intermediario del catabolismo de los globósidos y gangliósidos, lo que origina una acumulación de estos glucolípidos en los tejidos.
Esta patología fue descrita inicialmente como una variante de la enfermedad de Tay-Sachs, relacionada con el metabolismo de los gangliósidos, pero esta presenta también la acumulación de globósidos en las vísceras. Es un desorden hereditario con patrones autosómicos recesivos que destruye progresivamente neuronas y médula espinal.
Tiene que ver con la ausencia de las formas A y B de la enzima β-N-acetil hexosaminidasa debido a mutaciones en el gen HEXB. Estas enzimas son las encargadas de uno de los pasos de degradación de algunos glucoesfingolípidos.
- Bieberich, E. (2004). Integration of glycosphingolipid metabolism and cell-fate decisions in cancer and stem cells: Review and Hypothesis. Glycoconjugate Journal, 21, 315–327.
- Brady, R., Gal, A., Bradley, R., Martensson, E., Warshaw, A., & Laster, L. (1967). Enzymatic Defect in Fabry’s Disease. The New England Journal of Medicine, 276(21), 1163–1167.
- D’Angelo, G., Capasso, S., Sticco, L., & Russo, D. (2013). Glycosphingolipids: synthesis and functions. The FEBS Journal, 280, 6338–6353.
- Eto, Y., & Suzuki, K. (1971). Brain sphingoglycolipids in Krabbe’s globoid cell leukodystrophy. Journal of Neurochemistry, I(1966).
- Jones, D. H., Lingwood, C. A., Barber, K. R., & Grant, C. W. M. (1997). Globoside as a Membrane Receptor : A Consideration of Oligosaccharide Communication with the Hydrophobic Domain †. Biochemistry, 31(97), 8539–8547.
- Merrill, A. H. (2011). Sphingolipid and glycosphingolipid metabolic pathways in the era of sphingolipidomics. Chemical Reviews, 111(10), 6387–6422.
- Park, S., Kwak, C., Shayman, J. A., & Hoe, J. (2012). Globoside promotes activation of ERK by interaction with the epidermal growth factor receptor. Biochimica et Biophysica Acta, 1820(7), 1141–1148.
- U.S. Department of Health & Human Services (2008). Genetics Home Reference Sandhoff disease. Retrieved from www.ghr.nlm.nih.gov/condition/sandhoff-disease#definition
- Spence, M., Ripley, B., Embil, J., & Tibbles, J. (1974). A New Variant of Sandhoff’s Disease. Pediat. Res., 8, 628–637.
- Tatematsu, M., Imaida, K., Ito, N., Togari, H., Suzuki, Y., & Ogiu, T. (1981). Sandhoff Disease. Acta Pathol. Jpn, 31(3), 503–512.
- Traversier, M., Gaslondes, T., Milesi, S., Michel, S., & Delannay, E. (2018). Polar lipids in cosmetics: recent trends in extraction, separation, analysis and main applications. Phytochem Rev, 7, 1–32.
- Yamakawa, T., Yokoyama, S., & Kiso, N. (1962). Structure of Main Globoside of Human Erythrocytes. The Journal of Biochemistry, 52(3).