Rinoceronte lanudo: características, hábitat, causas, extinción
El rinoceronte lanudo (Coelodonta antiquitatis) es una especie de rinoceronte extinta que habitó gran parte de Europa y Asia durante el último período glacial. Junto con otras especies extintas de rinocerontes y las especies actuales forman parte del orden Perissodactyla y la familia Rinocerontidae. Fue descrita por Blumenbach en 1799, siendo Coelodonta antiquitatis la especie tipo del género y la más reciente en la escala temporal.
Cuando se descubrieron, los investigadores no podían creer que existieran rinocerontes que vivieran en condiciones con tan bajas temperaturas y, a partir de esto, surgieron muchas hipótesis (que más tarde fueron refutadas) para explicar su presencia en dichas zonas.

El rinoceronte lanudo forma parte de un conjunto de mamíferos de gran talla conocidos como mamíferos de la estepa del mamut o complejo faunístico “Mammuthus-Coelodonta”. C. antiquitatis constituía el segundo mamífero más grande después del mamut en el norte de Eurasia.
Estos animales están muy bien representados en la pintura rupestre al igual que otras especies de mamíferos de Pleistoceno, por lo que son considerados elementos típicos de la megafauna de este período.
Debido a que son pocos los animales que se han descubierto con tejidos blandos conservados, la información concerniente a su biología es escasa y gran parte de la información señalada se basa en analogías con especies de rinocerontes vivientes.
El pelaje que los recubría era abundante y de coloración parda. El comportamiento alimenticio es muy similar al de los rinocerontes actuales y estas evidencias están apoyadas en reconstrucciones paleo-climáticas, análisis de polen y modelos biométricos realizados a partir del cráneo.
La extinción de este mamífero en Europa coincide con un evento extendido de bajas temperaturas conocido como “dryas antiguo”. Aunque otros autores indican que la desaparición se debe a la expansión de los bosques como producto del cambio climático. Por otro lado, la extinción de las últimas poblaciones en Siberia se atribuye a un periodo de calentamiento conocido como Bølling-Allerød.
Índice del artículo
- 1 Características del rinoceronte lanudo
- 2 Taxonomía
- 3 Hábitat y distribución
- 4 Extinción y causas
- 5 Alimentación
- 6 Reproducción
- 7 Referencias
Eran animales de gran tamaño y de cuerpo robusto, incluso más grandes que los rinocerontes blancos.
Poseía extremidades cortas y un pelaje o lana abundante, además de una piel gruesa que lo aislaba térmicamente para sobrevivir en ambientes de tundra y estepa característicos de las regiones de altas latitudes durante las glaciaciones del pleistoceno.
La longitud total de estos grandes mamíferos oscilaba entre 3,5 y 3,8 metros en los machos y de 3, 2 a 3,6 metros en las hembras, con una altura que podía alcanzar los dos metros y de 1,6 metros a nivel de los hombros. El peso de estos animales era superior a las tres toneladas en los machos y cerca de dos toneladas en las hembras.
En contraste con los rinocerontes actuales, las orejas del rinoceronte lanudo eran más estrechas y la cola considerablemente más corta. Estas características reflejan la regla ecológica de Allen como una adaptación al clima frío.
El labio superior es corto y ancho similar al del rinoceronte blanco, siendo un indicativo adicional de su alimentación a base de pastos y cereales. En el siguiente video se puede observar cómo podría ser esta especie:
Como los rinocerontes actuales, los cuernos de los rinocerontes lanudos están formados por filamentos o fibras lamelares queratinizadas dispuestas paralelamente a lo largo del eje longitudinal. Estas fibras se empaquetan dentro de una matriz amorfa melanizada de polifasequeratina.
C. antiquitatis poseía dos cuernos al igual que el rinoceronte blanco Ceratotherium simum. Presentaban un cuerno nasal muy desarrollado con una longitud que oscila entre 70 cm y 1 metro de largo en los machos y de 60 cm a 70 cm en las hembras y un segundo cuerno frontal entre los ojos mucho más corto.
Los rasgos característicos de estos cuernos son la presencia de bandas transversales más o menos regulares con un par de capas, una clara y densa y una oscura y menos densa atribuidas al crecimiento anual. El cuerno nasal era aplanado lateralmente y ligeramente curvado.
El género Coelodonta, con todos sus representantes extintos, está constituido según el registro fósil por cuatro especies bien diferenciadas.
Coelodonta thibetana que constituye la especie más ancestral que hábito las mesetas del Tíbet, C. nihowanensis originaria del norte de China, C. tologoijensis también del norte de China y norte de Europa y C. antiquitatis la especie más ampliamente extendida.
Los parientes vivos más cercanos del rinoceronte lanudo son los rinocerontes de Sumatra Dicerorhinus sumatrensis.
Coelodonta antiquitatis también ha sido referida como Rhinocerus tichorhinus.

Según los hallazgos fósiles, Coelodonta antiquitatis habitó ampliamente el continente europeo y Asia. Sus fósiles se han encontrado en el área que abarca desde las Islas Británicas en el oeste hasta Chukotka y Kamchatka en el este de Rusia.
Se han hecho registros en prácticamente toda Europa occidental y central, en Europa oriental al norte del Cáucaso, Siberia occidental y el norte de Kazajstán, Siberia oriental y nororiental, norte de Mongolia y el norte y noreste de China. También existen reportes para la península escandinava y algunas regiones del ártico.
Por otro lado, existen registros no verificados de esta especie en Corea, cuya existencia es probable debido a la existencia de rinocerontes lanudos en China.
Los paisajes dominantes en donde se desenvolvían estos grandes mamíferos eran predominantemente conocidos como tundra estepa o estepa del mamut, propios de climas muy fríos y secos y característicos de los períodos glaciales. La vegetación era principalmente herbácea esteparia con escasa vegetación arbórea.

Esta especie no se extendió hasta América del Norte como otras especies de grandes herbívoros de la época, a pesar de que estuvo presente en regiones cercanas al estrecho de Bering cuando el puente se elevaba por encima del nivel de mar.
Probablemente las condiciones húmedas y pantanosas le impidieron a este animal de patas cortas y pelo largo atravesar este enlace continental.
Por otro lado, mamíferos como el mamut lanudo tenían tolerancias ecológicas superiores a las que presentaba el rinoceronte lanudo una razón adicional que explica porque estos rinocerontes no colonizaron el nuevo mundo.
La extinción de estos animales probablemente se debe a la sinergia de varios factores como el cambio climático, las interacciones con el hombre, la disponibilidad de recursos y la competencia por estos y la aparición de enfermedades.
La desaparición de esta especie en su área de distribución original ocurrió de manera escalonada con retraso en algunas regiones de hasta varios miles de años. La pérdida de pastizales cruciales para la alimentación fue un factor determinante.
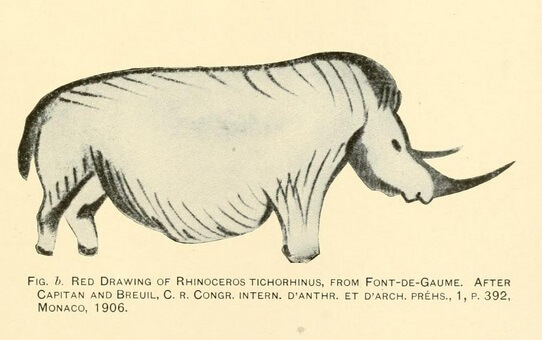
Las últimas poblaciones en extinguirse fueron las distribuidas más al norte, al noreste de Europa (Rusia Europea y los estados Bálticos y la parte central y el norte de Siberia).
Las dataciones con C14 indican que los rinocerontes desaparecieron hace unos 14000 -13600 años en Europa. Mientras, en Asia existen dataciones entre los 14400-12800 años. Las dataciones más recientes corresponden con registros hechos en Siberia con entre 13170 y 10770 años.
Es muy probable que el factor nival desempeñara un papel fundamental en la extinción del rinoceronte lanudo.
El aumento de la temperatura y las precipitaciones al final de Pleistoceno y principios del Holoceno (hace unos 10.000 años) provocó la fragmentación del tipo de hábitat de estepa ampliamente extendido en Eurasia. Seguido a esto, los ambientes de estepa-tundra fueron sustituidos por otras coberturas vegetales más arbustivas y boscosas.
Adicionalmente, el calentamiento y aumento de la humedad probablemente provocaron que en la época de invierno las capas de nieve aumentaran en altura. Esto tuvo como consecuencia serios impedimentos en la movilización del rinoceronte lanudo y originó además limitaciones en la búsqueda de alimentos bajo la nieve.
Por otro lado, como consecuencia del cambio climático, surgieron trampas naturales como cavidades después del derretimiento del hielo, pantanos y arroyos irregulares.
Debido a que el rinoceronte lanudo presentaba extremidades muy cortas y eran pesados, estos cambios en el terreno eran potencialmente mortales. Varios de los fósiles encontrados, parecen haber muerto bajo estas condiciones particulares.
Muchas teorías señalan que el efecto de la caza excesiva de grandes mamíferos por parte del hombre pudo influenciar el decremento de las poblaciones de estos animales y su posterior desaparición. En el arte rupestre han sido identificados muchos retratos del rinoceronte lanudo, muy fieles a las características que hoy conocemos.
Probablemente estos retratos se deban a que esta especie tuvo cierto grado de importancia para la subsistencia del hombre.
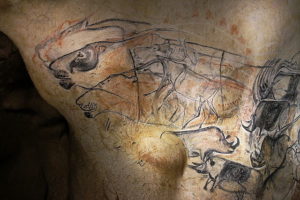
Otra de las teorías que explican la desaparición de una gran cantidad de especies simultáneamente durante el pleistoceno es conocida como la “súper enfermedad”. Sin embargo, hasta el momento no se conoce ninguna enfermedad que haya provocado la desaparición de grupos animales de diversas familias y órdenes e incluso infraclases.
Con base a la morfología de los dientes y a las características similares a los rinocerontes actuales se ha concluido que esta especie de alimentaba de vegetación herbácea y de pastizales. Estos animales estaban adaptados para el consumo de vegetación herbácea de crecimiento lento en ambientes fríos y secos con nevadas mínimas.
A partir de los registros fósiles en gran parte de Europa y Asia, se ha precisado que no era un animal migratorio en comparación con otras especies de megaherbívoros del Pleistoceno como el mamut lanudo.
Algunos restos de plantas extraídos de la boca de rinocerontes lanudos bien conservados en el hielo contenían restos de pastos. Además de esto, contenían ramas de coníferas (Picea y Larix) y árboles caducifolios como Salix.
Análisis palinológicos han determinado la presencia de diversas especies de gramíneas, Artemisia, Betula, Alnus, helechos y musgos.

Como el mamut lanudo, C. antiquitatis probablemente se alimentaba de pastos y juncias la mayor parte del año. Sin embargo, al no ser un animal migratorio es posible que durante la temporada de nevadas y precipitaciones su alimentación variara.
Los análisis de isótopos estables en los cuernos junto con aquellos presentes en el suelo congelado (permafrost) alrededor de los fósiles, revelan que las diferencias en la composición de las capas de tejido cartilaginoso del cuerno se deben principalmente a un cambio estacional en la dieta. Esto también ocurre con los cuernos de algunos mamíferos actuales.
Las zonas oscuras y menos densas del cuerno se asocian con una alimentación a base de hierbas y pastos superficiales dado el contenido positivo de C13 y N15, coincidiendo con la típica alimentación de verano. Por otro lado, las zonas más claras y menos densas se asocian con alimentación a partir de herbáceas y leñosas durante el invierno.
Esta especie presentaba un gran dimorfismo sexual superior al de las especies vivientes. Los cuernos de los machos eran bastante desarrollados, por lo que se cree tenían una función de atracción sobre las hembras además de emplearlos en las luchas contra otros machos y como medida defensiva ante los depredadores.

Es probable que las características reproductivas fuesen similares a las de los rinocerontes actuales. Estos animales tenían tasas reproductivas bajas. El descubrimiento de hembras bien conservadas con una ubre de dos pezones indica que una hembra probablemente solo paría a una cría y excepcionalmente a dos.
Por analogía con las especies de rinocerontes actuales, se indica que un hembra podía quedar preñada cada dos o tres años o incluso más. No se conoce si existía una época reproductiva específica a lo largo del año, pues la mayoría de los hallazgos de rinocerontes lanudos o partes grandes bien conservadas pertenecen a hembras.
Es posible que, debido al tamaño y forma del cuerno de estos rinocerontes, en la época de bajas temperaturas y escasa disponibilidad de alimento estos animales se valieran de sus enormes cuernos para cavar en la nieve y exponer la vegetación de la cual se alimentaba.

Se sabe por datos indirectos, así como la presencia de otros mamíferos, que las capas de nieve que se formaban en los hábitats de estos animales no superaban los 30 cm, lo que favorecía su desplazamiento entre ellas.
Aquellos hábitats que presentaban nieve más abundante constituía un impedimento para la movilidad de estos animales y probablemente evitaron su dispersión al continente americano.
La existencia de cráneos de rinocerontes lanudos con algunos traumatismos leves y severos, indican que muy probablemente estos animales hayan sido fuertemente territoriales.
Al igual que el caso de los rinocerontes actuales, las luchas entre rinocerontes solían ser frecuentes y en raros casos provocaban lesiones graves en el cráneo. Quizás, debido a los cambios en la disponibilidad de recursos en el invierno, las relaciones entre individuos aumentaban en agresividad, producto de la competencia intraespecífica.
- Boeskorov, G. G. (2012). Some specific morphological and ecological features of the fossil woolly rhinoceros (Coelodonta antiquitatis Blumenbach 1799). Biology bulletin, 39(8), 692-707.
- Deng, T., Wang, X., Fortelius, M., Li, Q., Wang, Y., Tseng, Z. J., … & Xie, G. (2011). Out of Tibet: Pliocene woolly rhino suggests high-plateau origin of Ice Age megaherbivores. Science, 333(6047), 1285-1288.
- Fortelius, M. (1983). The morphology and paleobiological significance of the horns of Coelodonta antiquitatis (Mammalia: Rhinocerotidae). Journal of Vertebrate Paleontology, 3(2), 125-135.
- Garutt, N. (1997). Traumatic skull damages in the woolly rhinoceros, Coelodonta antiquitatis Blumenbach, 1799. Cranium, 14(1), 37-46.