Ácidos grasos: estructura, tipos, funciones, biosíntesis
Los ácidos grasos son macromoléculas orgánicas derivadas de hidrocarburos, que están compuestas por largas cadenas de átomos de carbono e hidrógeno que tienen carácter hidrofóbico (son liposolubles) y son la base estructural de las grasas y los lípidos.
Son moléculas muy diversas que se distinguen unas de otras por la longitud de sus cadenas hidrocarbonadas y la presencia, el número, la posición y/o configuración de sus dobles enlaces.
En los lípidos de animales, plantas, hongos y microorganismos como las bacterias y las levaduras, se han descrito más de 100 clases de ácidos grasos diferentes y se considera que son especie y tejido específicos en la mayor parte de los seres vivos.
Los aceites y las grasas que el hombre consume diariamente sean de origen animal o vegetal, están compuestas mayoritariamente por ácidos grasos.
Índice del artículo
- 1 Introducción
- 2 Estructura
- 3 Clasificación
- 4 Funciones
- 5 Biosíntesis
- 6 Propiedades de los ácidos grasos
- 7 Nomenclatura
- 8 Referencias

Las moléculas de ácidos grasos desempeñan importantes funciones a nivel celular, lo que los hace componentes esenciales y, puesto que algunos de ellos no pueden ser sintetizados por los animales, estos deben obtenerlos de la dieta.
Los ácidos grasos son poco comunes como especies libres en el citosol celular, por lo que generalmente se encuentran formando parte de otros conjugados moleculares como:
– Lípidos, en las membranas biológicas.
– Triglicéridos o ésteres de ácidos grasos, que sirven de reserva en plantas y animales.
– Ceras, que son ésteres sólidos de ácidos grasos de cadena larga y alcoholes.
– Otras sustancias similares.
En los animales, los ácidos grasos se almacenan en el citoplasma de las células como pequeñas gotas de grasa compuestas por un complejo denominado triacilglicerol, que no es más que una molécula de glicerol a la cual se ha unido, en cada uno de sus átomos de carbono, una cadena de ácido graso por enlaces de tipo éster.
Mientras que las bacterias poseen ácidos grasos cortos y usualmente monoinsaturados, en la naturaleza es común hallar ácidos grasos cuyas cadenas poseen números pares de átomos de carbono, usualmente entre 14 y 24, saturados, monoinsaturados o poliinsaturados.
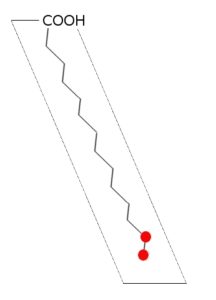
Los ácidos grasos son moléculas anfipáticas, es decir, que poseen dos regiones químicamente definidas: una región polar hidrofílica y una región apolar hidrofóbica.
La región hidrofóbica está conformada por una larga cadena hidrocarbonada que, en términos químicos, es poco reactiva. La región hidrofílica, por otra parte, está compuesta por un grupo carboxilo (-COOH) terminal, que se comporta como un ácido.
Este grupo carboxilo terminal o ácido carboxílico se ioniza en solución, es altamente reactivo (químicamente hablando) y es muy hidrófilo, por lo que representa un sitio de unión covalente entre el ácido graso y otras moléculas.
La longitud de las cadenas hidrocarbonadas de los ácidos grasos usualmente posee números pares de átomos de carbono, y ello está estrechamente relacionado con el proceso biosintético por el cual estos son producidos, puesto que su crecimiento ocurre de a pares de carbonos.
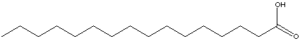
Los ácidos grasos más comunes poseen cadenas de entre 16 y 18 átomos de carbono y en los animales, estas cadenas no están ramificadas.
Los ácidos grasos se clasifican en dos grandes grupos según la naturaleza de los enlaces que los componen, es decir, según la presencia de enlaces sencillos o enlaces dobles entre los átomos de carbono de sus cadenas hidrocarbonadas.
Así, existen los ácidos grasos saturados e insaturados.
– Los ácidos grasos saturados poseen únicamente enlaces simples carbono – carbono y todos sus átomos de carbono están “saturados” o unidos a moléculas de hidrógeno.
– Los ácidos grasos insaturados presentan uno o más dobles enlaces carbono – carbono y no todos estos están unidos a un átomo de hidrógeno.
Los ácidos grasos insaturados también se dividen de acuerdo al número de insaturaciones (dobles enlaces) en monoinsaturados, aquellos con un solo doble enlace, y poliinsaturados, los que tienen más de uno.
Usualmente tienen entre 4 y 26 átomos de carbono unidos por enlaces sencillos. Su punto de fusión es directamente proporcional al largo de la cadena, es decir, a su peso molecular.
Los ácidos grasos que poseen entre 4 y 8 carbonos son líquidos a 25°C y son los que componen los aceites comestibles, mientras que los que tienen más de 10 átomos de carbono son sólidos.
Entre los más comunes se encuentra el ácido láurico, que es abundante en aceites de semilla de palma y de coco; el ácido palmítico, que se encuentra en la palma, el cacao y la manteca de porcinos, y el ácido esteárico, que está en el cacao y los aceites hidrogenados.
Son ácidos grasos con mucha más estabilidad que los ácidos grasos insaturados, especialmente frente a oxidación, al menos en condiciones fisiológicas.
Gracias a que los enlaces sencillos carbono – carbono pueden rotar con libertad, los ácidos grasos saturados son moléculas muy flexibles, aunque el impedimento estérico hace que la estructura completamente extendida sea la más estable energéticamente hablando.
Estos ácidos grasos son altamente reactivos y propensos a la saturación y oxidación. Son comunes en plantas y en organismos marinos. Los que poseen un solo doble enlace se conocen como monoinsaturados o monoenoicos, mientras que los que poseen más de dos se conocen como polienoicos o poliinsaturados.
La presencia de dobles enlaces es habitual entre los átomos de carbono entre las posiciones 9 y 10, pero esto no quiere decir que no se encuentren ácidos grasos monoinsaturados con una insaturación en otra posición.
A diferencia de los saturados, los ácidos grasos insaturados se enumeran no a partir del grupo carboxilo terminal, sino de acuerdo con la posición del primer enlace doble C – C. Así, estos se dividen en dos grupos, los ácidos omega-6 o ω6 y los omega-3 o ω3.
Los ácidos omega-6 tienen el primer doble enlace en el carbono número 6 y los ácidos omega-3 lo tienen en el carbono número 3. La denominación ω se da por el doble enlace más cercano al grupo metilo final.
Los dobles enlaces pueden hallarse, además, en dos configuraciones geométricas conocidas como “cis” y “trans”.
La mayor parte de los ácidos grasos insaturados naturales poseen configuración “cis” y los dobles enlaces de los ácidos grasos presentes en las grasas comerciales (hidrogenadas) se encuentran en “trans”.
En los ácidos grasos poliinsaturados, dos dobles enlaces están usualmente separados entre sí al menos por un grupo metilo, es decir, un átomo de carbono unido a dos átomos de hidrógeno.
Los ácidos grasos tienen múltiples funciones en los organismos vivos y, como se comentó anteriormente, una de sus funciones esenciales en como parte imprescindible de los lípidos, que son los componentes principales de las membranas biológicas y una de las tres biomoléculas más abundantes en los organismos vivos en conjunto con las proteínas y los carbohidratos.
Son, además, excelentes sustratos energéticos gracias a los cuales se obtienen grandes cantidades de energía en forma de ATP y otros metabolitos intermediarios.
En vista de que los animales, por ejemplo, no son capaces de almacenar carbohidratos, los ácidos grasos representan la principal fuente de almacenamiento energético que proviene de la oxidación de los azúcares consumidos en exceso.
Los ácidos grasos saturados de cadena corta en el colon participan en la estimulación de la absorción de agua e iones de sodio, cloruro y bicarbonato; además, tienen funciones en la producción de moco, en la proliferación de los colonocitos (células del colon), etc.
Los ácidos grasos insaturados son especialmente abundantes en los aceites comestibles de procedencia vegetal, que son importantes en la dieta de todos los seres humanos.

Otros participan como ligandos de algunas proteínas con actividades enzimáticas, por lo que son importantes con respecto a sus efectos en el metabolismo energético de las células donde se encuentran.
La degradación de ácidos grasos se conoce como β-oxidación y ocurre en la mitocondria de las células eucariotas. La biosíntesis, por el contrario, ocurre en el citosol de las células animales y en los cloroplastos (orgánulos fotosintéticos) de las células vegetales.
Es un proceso dependiente de acetil-CoA, malonil-CoA y NADPH, ocurre en todos los organismos vivos y en los animales “superiores” como los mamíferos. Por ejemplo, es muy importante en los tejidos hepáticos y adiposos, así como en las glándulas mamarias.
El NADPH empleado para esta ruta es producto, principalmente, de las reacciones de oxidación NADP-dependientes de la ruta de las pentosas fosfato, mientras que el acetil-CoA puede provenir de diferentes fuentes, por ejemplo, de la descarboxilación oxidativa del piruvato, del ciclo de Krebs y de la β-oxidación de ácidos grasos.
La ruta de biosíntesis, al igual que la de β-oxidación, está altamente regulada en todas las células por efectores alostéricos y modificaciones covalentes de enzimas que participan en la regulación.
La ruta comienza con la formación de un intermediario metabólico conocido como malonil-CoA a partir de una molécula de acetil-CoA y es catalizada por una enzima multifuncional llamada acetil-CoA carboxilasa.
Esta reacción es una reacción de adición de una molécula de carboxilo (-COOH, carboxilación) dependiente de biotina y ocurre en dos pasos:
- Primero se da la transferencia, dependiente de ATP, de un carboxilo derivado del bicarbonato (HCO3-) a una molécula de biotina que se encuentra como grupo prostético (no proteico) asociado a la acetil-CoA carboxilasa.
- Posteriormente, el CO2 es transferido al acetil-coA y se produce malonil-coA.
En los animales, la formación de las cadenas carbohidratadas de los ácidos grasos ocurre ulteriormente a través de reacciones de condensación secuenciales catalizadas por una enzima multimérica y multifuncional que se conoce como la ácido graso sintasa.
Esta enzima cataliza la condensación de una unidad de acetil-CoA y múltiples moléculas de malonil-CoA que son producidas a partir de la reacción de la acetil-CoA carboxilasa, proceso durante el cual se libera una molécula de CO2 por cada malonil-CoA que se añade.
Los ácidos grasos en crecimiento se encuentran esterificados a una proteína denominada “proteína portadora de acilo” o ACP, que forma tioésteres con los grupos acilo. En E. coli esta proteína es un polipéptido de 10 kDa, pero en los animales es parte del complejo ácido graso sintasa.
La ruptura de estos enlaces tioéster libera grandes cantidades de energía, que hace posible, termodinámicamente hablando, la ocurrencia de los pasos de condensación en la ruta biosintética.
Complejo ácido graso sintasa
En bacterias, la actividad ácido graso sintasa corresponde en realidad a seis enzimas independientes que emplean acetil-coA y malonil-coA para formar los ácidos grasos y con las que se asocian seis actividades enzimáticas distintas.
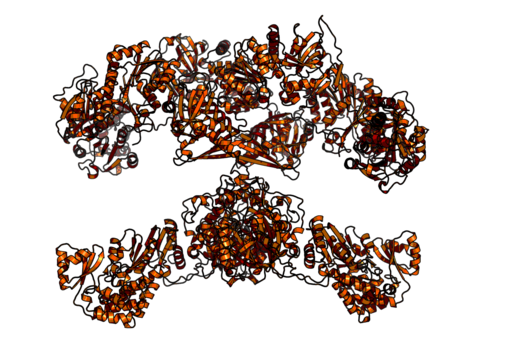
En los mamíferos, por el contrario, la ácido graso sintasa es un complejo enzimático homodimérico multifuncional de alrededor de 500 kDa de peso molecular, que tiene seis actividades catalíticas diferentes y con la que se asocia la proteína portadora de acilo.
Paso 1: Reacción cebadora
Los grupos tiol en los residuos de cisteína responsables de la unión de los intermediarios metabólicos a la enzima ACP deben ser cargados, antes del comienzo de la síntesis, con los grupos acilo necesarios.
Para ello, el grupo acetilo del acetil-coA es transferido al grupo tiol (-SH) de uno de los residuos de cisteína de la subunidad ACP de la ácido graso sintasa. Esta reacción es catalizada por la subunidad ACP-acil-transferasa.
El grupo acetilo es luego transferido de la ACP hacia otro residuo de cisteína en el sitio catalítico de otra subunidad enzimática del complejo que se conoce como β-cetoacil-ACP-sintasa. Así, el complejo enzimático queda “cebado” para comenzar la síntesis.
Paso 2: Transferencia de unidades de malonil-CoA
El malonil-CoA que es producido por la acetil-CoA carboxilasa es transferido al grupo tiol en la ACP y durante esta reacción se pierde la porción CoA. La reacción es catalizada por la subunidad malonil-ACP-transferasa del complejo ácido graso sintasa, que produce entonces malonil-ACP.
Durante este proceso, el grupo malonilo se encuentra unido a la ACP y a la β-cetoacil-ACP-sintasa a través de un enlace éster y otro sulfidrilo, respectivamente.
Paso 3: Condensación
La enzima β-cetoacil-ACP-sintasa cataliza la transferencia del grupo acetilo que se le unió en el paso de “cebado” hacia el carbono 2 del grupo malonilo que, en el paso anterior, fue transferido a la ACP.
Durante esta reacción se libera una molécula de CO2 que proviene del malonilo, que corresponde al CO2 aportado por el bicarbonato en la reacción de carboxilación de la acetil-CoA carboxilasa. Se produce, entonces, acetoacetil-ACP.
Paso 4: Reducción
La subunidad β-cetoacil-ACP-reductasa cataliza la reducción del acetoacetil-ACP dependiente de NADPH, con lo que se forma D-β-hidroxibutiril-ACP.
Paso 5: Deshidratación
En este paso se forma el trans-α, β-acil-ACP ó el ∆2-insaturado-acil-ACP (cratonil-ACP), producto de la deshidratación del D-β-hidroxibutiril-ACP por acción de la subunidad enoil-ACP-hidratasa.
Más adelante, el cratonil-ACP se reduce a butiril-ACP por una reacción dependiente de NADPH catalizada por la subunidad enoil-ACP-reductasa. Con esta reacción se completa el primero de siete ciclos que se necesitan para producir palmitoil-ACP, que es precursor de casi todos los ácidos grasos.
¿Cómo siguen las reacciones de condensación posteriores?
El grupo butirilo se transfiere del ACP al grupo tiol de un residuo de cisteína en la β-cetoacil-ACP-sintasa, con lo que la ACP es capaz de aceptar otro grupo malonilo a partir el malonil-CoA.
De esta manera, la reacción que sucede es la condensación de malonil-ACP con buturil-β-cetoacil-ACP-sintasa, que da lugar al β-cetohexanoil-ACP + CO2.
El palmitoil-ACP que surge a partir de los pasos subsecuentes (luego de la adición de 5 unidades de malonilo más) puede liberarse como ácido palmítico libre gracias a la actividad de la enzima tioesterasa, puede transferirse al CoA o incorporarse al ácido fosfatídico para la ruta de síntesis de fosfolípidos y triacilglicéridos.
La ácido graso sintasa de la mayoría de los organismos se detiene en la síntesis de palmitoil-ACP, dado que el sitio catalítico de la subunidad β-cetoacil-ACP-sintasa tiene una configuración en la cual solo pueden acomodarse ácidos grasos de esa longitud.
Estos son relativamente comunes en los organismos marinos y también son sintetizados por un complejo ácido graso sintasa. No obstante, la reacción de “cebado” se da con una molécula más larga, el propionil-ACP, de tres átomos de carbono.
El ácido palmítico, como se comentó, sirve de precursor para muchos ácidos grasos saturados e insaturados de cadenas más largas. El proceso de “alargamiento” de los ácidos grasos ocurre en la mitocondria, mientras que la introducción de insaturaciones ocurre esencialmente en el retículo endoplásmico.
Muchos organismos convierten sus ácidos grasos saturados en insaturados como una adaptación a bajas temperaturas medioambientales, puesto que ello les permite mantener el punto de fusión de los lípidos por debajo de la temperatura ambiente.
Muchas de las propiedades de los ácidos grasos dependen de su longitud de cadena y de la presencia y número de insaturaciones:
– Los ácidos grasos insaturados tienen puntos de fusión más bajos que los ácidos grasos saturados de la misma longitud.
– La longitud de los ácidos grasos (el número de átomos de carbono) es inversamente proporcional a la fluidez o flexibilidad de la molécula, es decir, las moléculas más “cortas” son más fluidas y viceversa.
Por lo general, las sustancias grasas fluidas están compuestas por ácidos grasos de cadenas cortas y con presencia de insaturaciones.
Las plantas poseen abundante cantidad de ácidos grasos insaturados, así como los animales que viven a muy bajas temperaturas, pues estos, como componentes de los lípidos presentes en las membranas celulares, les otorgan mayor fluidez en estas condiciones.
En condiciones fisiológicas, la presencia de un doble enlace en la cadena hidrocarbonada de un ácido graso causa una curvatura de unos 30°, lo que hace que estas moléculas ocupen un espacio mayor y disminuyan la fuerza de sus interacciones de van der Waals.
La presencia de dobles enlaces en los ácidos grasos asociados con moléculas lipídicas tiene efectos directos en el grado de “empaquetamiento” que estos puedan tener en las membranas a las que pertenecen y con ello tienen efectos también sobre las proteínas membranales.
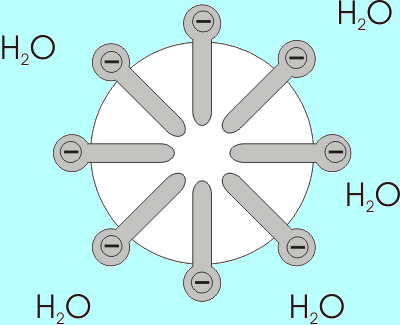
La solubilidad de los ácidos grasos disminuye a medida que aumenta la longitud de su cadena, por lo que son inversamente proporcionales. En mezclas acuosas y lipídicas, los ácidos grasos se asocian en estructuras conocidas como micelas.
Una micela es una estructura en la cual se “encierran” las cadenas alifáticas de los ácidos grasos, “expulsando” así todas las moléculas de agua y en cuya superficie se encuentran los grupos carboxilos.
La nomenclatura de los ácidos grasos puede ser un tanto compleja, especialmente si uno se refiere a los nombres comunes que estos reciben, que a menudo se relacionan con alguna propiedad fisicoquímica, con el lugar donde se encuentran u otras características.
Muchos autores consideran que como gracias al grupo carboxilo terminal estas moléculas están ionizadas a pH fisiológico, uno debería referirse a ellas como “carboxilatos” empleando para ello la terminación “ato”.
De acuerdo con el sistema IUPAC, la enumeración de los átomos de carbono de un ácido graso se hace a partir del grupo carboxilo en el extremo polar de la molécula y los dos primeros átomos de carbono unidos a este grupo se denominan α y β, respectivamente. El metilo terminal de la cadena contiene el átomo de carbono ω.
Por lo general, en la nomenclatura sistemática se les otorga el nombre del hidrocarburo “parental” (el hidrocarburo con el mismo número de átomos de carbono) y se sustituye su terminación “o” por “oico”, si se trata de un ácido graso insaturado, se añade la terminación “enoico”.
Considérese, por ejemplo, el caso de un ácido graso de 18 átomos de carbono (C18):
– Puesto que el hidrocarburo con la misma cantidad de átomos de carbono se conoce como octadecano, el ácido saturado se denomina “ácido octadecanoico” o bien “octadecanoato” y su nombre común es ácido esteárico.
– Si tiene un doble enlace entre algún par de átomos de carbono en su estructura, se conoce como “ácido octadecenoico”
– Si tiene dos enlaces dobles c – c, entonces se denomina “ácido octadecadienoico” y si tiene tres “ácido octadecatrienoico”.
Si se quiere resumir la nomenclatura, entonces de emplea 18:0 para el ácido graso de 18 carbonos y ningún doble enlace (saturado) y, dependiendo del grado de insaturación, entonces en vez de cero se escribe 18:1 para una molécula con una insaturación, 18:2 para una con dos insaturaciones y así sucesivamente.
Si se quiere especificar entre cuáles átomos de carbono se encuentran los dobles enlaces en los ácidos grasos insaturados, se emplea el símbolo ∆ con un superíndice numérico que indica el lugar de la insaturación y el prefijo “cis” o “trans”, dependiendo de la configuración de este.
- Badui, S. (2006). Química de los alimentos. (E. Quintanar, Ed.) (4ta ed.). México D.F.: Pearson Education.
- Garrett, R., & Grisham, C. (2010). Biochemistry (4th ed.). Boston, USA: Brooks/Cole. CENGAGE Learning.
- Mathews, C., van Holde, K., & Ahern, K. (2000). Biochemistry (3th ed.). San Francisco, California: Pearson.
- Murray, R., Bender, D., Botham, K., Kennelly, P., Rodwell, V., & Weil, P. (2009). Harper’s Illustrated Biochemistry (28th ed.). McGraw-Hill Medical.
- Nelson, D. L., & Cox, M. M. (2009). Lehninger Principios de Bioquímica. Ediciones Omega (5th ed.).
- Rawn, J. D. (1998). Biochemistry. Burlington, Massachusetts: Neil Patterson Publishers.
- Tvrzicka, E., Kremmyda, L., Stankova, B., & Zak, A. (2011). Fatty acids as Biocompounds: Their Role in Human Metabolism, Health and Disease- A Review. Part 1: Classification, Dietary Sources and Biological Functions. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub, 155(2), 117–130.