Esfingomielina: estructura, funciones, síntesis y metabolismo
La esfingomielina es el esfingolípido más abundante en los tejidos animales: se ha comprobado su presencia en todas las membranas celulares estudiadas hasta la fecha. Posee similitudes estructurales con la fosfatidilcolina en cuanto al grupo de la cabeza polar, por lo que también se clasifica como fosfolípido (fosfoesfingolípido).
En la década de los años 1880, el científico Johann Thudichum aisló un componente lipídico éter-soluble a partir de tejido cerebral y lo bautizó como esfingomielina. Más tarde, en el año 1927, la estructura de este esfingolípido fue reportada como N-acil-esfingosina-1-fosfocolina.
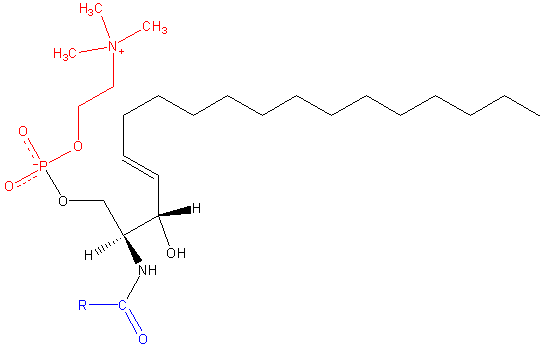
Al igual que los demás esfingolípidos, la esfingomielina tiene funciones tanto estructurales como de señalización celular, y es especialmente abundante en tejidos nerviosos, concretamente en la mielina, una vaina que recubre y aísla los axones de ciertas neuronas.
Su distribución ha sido estudiada a través de experimentos de fraccionamiento subcelular y de degradación enzimática con esfingomielinasas, y los resultados indican que más de la mitad de la esfingomielina en las células eucariotas se encuentra en la membrana plasmática. No obstante, ello depende del tipo celular. En los fibroblastos, por ejemplo, representa casi el 90% de los lípidos totales.
La desregulación de los procesos de síntesis y metabolismo de este lípido conllevan al desarrollo de patologías complejas o lipidosis. Un ejemplo de estas es la enfermedad hereditaria de Niemann-Pick, caracterizada por hepatoesplenomegalia y disfunción neurológica progresiva.
Índice del artículo
La esfingomielina es una molécula anfipática compuesta por una cabeza polar y dos colas apolares. El grupo de cabeza polar es una molécula de fosfocolina, por lo que puede parecer similar al glicerofosfolípido fosfatidilcolina (PC). No obstante, existen diferencias sustanciales respecto a la región interfacial e hidrofóbica entre estas dos moléculas.
La base más común en una molécula de esfingomielina de mamíferos es la ceramida, compuesta por la esfingosina (1,3-dihidroxi-2-amino-4-octadeceno), que posee un doble enlace en trans entre los carbonos de las posiciones 4 y 5 de la cadena hidrocarbonada. Su derivado saturado, la esfinganina, también es común, pero se encuentra en menor proporción.
La longitud de las colas hidrofóbicas de la esfingomielina oscila entre 16 y 24 átomos de carbono y la composición de ácidos grasos varía dependiendo del tejido.
Las esfingomielinas de la materia blanca del cerebro humano, por ejemplo, poseen ácido nervónico, las de la materia gris contienen principalmente ácido esteárico, y la forma prevalente en las plaquetas es el araquidonato.
Generalmente existe una disparidad de longitud entre las dos cadenas de ácidos grasos de la esfingomielina, lo que parece favorecer fenómenos de “interdigitación” entre los hidrocarburos en monocapas opuestas. Esto proporciona a la membrana una estabilidad y propiedades particulares especiales respecto a otras membranas más pobres en este esfingolípido.
En la región interfacial de la molécula, la esfingomielina posee un grupo amida y un hidroxilo libre en el carbono 3, que pueden servir como donadores y aceptores de puentes de hidrógeno para enlaces intra- e intermoleculares, importantes en la definición de dominios laterales e interacción con diversos tipos de moléculas.
Los productos del metabolismo de la esfingosina -ceramida, esfingosina, esfingosina 1-fosfato y diacilglicerol-, son importantes efectores celulares y le dan un papel en múltiples funciones celulares, como la apoptosis, el desarrollo y envejecimiento, señalización celular, entre otros.
Gracias a la estructura tridimensional “cilíndrica” de la esfingomielina, este lípido puede formar dominios de membrana más compactos y ordenados, lo que tiene importantes implicaciones funcionales desde el punto de vista proteico, puesto que pueden establecer dominios específicos para algunas proteínas integrales de membrana.
En las “balsas” lipídicas y caveolas
Las balsas lipídicas, fases de membrana o micro dominios ordenados de esfingolípidos como la esfingomielina, algunos glicerofosfolípidos y colesterol, representan plataformas estables para la asociación de proteínas de membrana con diversas funciones (receptores, transportadores, etc.).
Las caveolas son invaginaciones de la membrana plasmática que reclutan proteínas con anclas GPI y también son ricas en esfingomielina.
En relación con el colesterol
El colesterol, debido a su rigidez estructural, afecta significativamente la estructura de las membranas celulares, especialmente en los aspectos relacionados con la fluidez, por lo que se considera un elemento esencial.
Gracias a que las esfingomielinas poseen tanto donadores como aceptores de puentes de hidrógeno, se cree que son capaces de formar interacciones más “estables” con las moléculas de colesterol. Por esto se dice que existe una correlación positiva entre los niveles de colesterol y esfingomielina en las membranas.
La síntesis de la esfingomielina ocurre en el complejo de Golgi, donde la ceramida transportada desde el retículo endoplasmático (ER) es modificada por la transferencia de una molécula fosfocolina proveniente de la fosfatidilcolina, con la liberación concomitante de una molécula de diacilglicerol. La reacción es catalizada por la SM sintasa (ceramida:fosfatidilcolina fosfocolina transferasa).
Existe también otra vía de producción de esfingomielina que puede ocurrir por transferencia de una fosfoetanolamina desde la fosfatidiletanolamina (PE) hacia la ceramida, con la posterior metilación de la fosfoetanolamina. Se piensa que esta puede ser particularmente importante en algunos tejidos nerviosos ricos en PE.
La esfingomielina sintasa se encuentra en el lado luminal de la membrana del complejo de Golgi, lo que coincide con la ubicación extra citoplasmática de la esfingomielina en la mayor parte de las células.
Debido a las características del grupo polar de la esfingomielina y a la aparente ausencia de translocasas específicas, la orientación topológica de este lípido depende de la enzima sintasa.
La degradación de la esfingomielina puede ocurrir tanto en la membrana plasmática como en los lisosomas. La hidrólisis lisosomal hasta ceramida y fosfocolina depende de la esfingomielinasa acídica, una glicoproteína lisosomal soluble cuya actividad tiene un pH óptimo alrededor de 4.5.
La hidrólisis en la membrana plasmática es catalizada por una esfingomielinasa que trabaja a pH 7.4 y que requiere de iones de magnesio o manganeso divalentes para su funcionamiento. Otras enzimas involucradas en el metabolismo y reciclaje de la esfingomielina se encuentran en diferentes organelas que se conectan entre sí a través de las vías de transporte vesicular.
- Barenholz, Y., & Thompson, T. E. (1999). Sphingomyelin : biophysical aspects. Chemistry and Physics of Lipids, 102, 29–34.
- Kanfer, J., & Hakomori, S. (1983). Sphingolipid Biochemistry. (D. Hanahan, Ed.), Handbook of Lipid Research 3 (1st ed.). Plenum Press.
- Koval, M., & Pagano, R. (1991). Intracellular transport and metabolism of sphingomyelin. Biochimic, 1082, 113–125.
- Lodish, H., Berk, A., Kaiser, C. A., Krieger, M., Bretscher, A., Ploegh, H., Martin, K. (2003). Molecular Cell Biology (5th ed.). Freeman, W. H. & Company.
- Millat, G., Chikh, K., Naureckiene, S., Sleat, D. E., Fensom, A. H., Higaki, K., … Vanier, M. T. (2001). Niemann-Pick Disease Type C : Spectrum of HE1 Mutations and Genotype / Phenotype Correlations in the NPC2 Group. Am. J. Hum. Genet., 69, 1013–1021.
- Ramstedt, B., & Slotte, P. (2002). Membrane properties of sphingomyelins. FEBS Letters, 531, 33–37.
- Slotte, P. (1999). Sphingomyelin – cholesterol interactions in biological and model membranes. Chemistry and Physics of Lipids, 102, 13–27.
- Vance, J. E., & Vance, D. E. (2008). Biochemistry of lipids, lipoproteins and membranes. In New Comprehensive Biochemistry Vol. 36 (4th ed.). Elsevier.