Aldosas: características, tipos, número de carbonos
Las aldosas son monosacáridos que poseen un grupo aldehído terminal. Son azúcares simples ya que no pueden ser hidrolizados para producir otros azúcares más simples. Poseen entre tres y siete átomos de carbono. Tal como las cetosas, las aldosas son azúcares polihidroxilados.
En la naturaleza, las aldosas más abundantes son arabinosa, galactosa, glucosa, manosa, ribosa y xilosa. En los organismos fotosintéticos, la biosíntesis de estos azúcares tiene lugar a partir de la fructosa-6-fosfato, un azúcar del ciclo de Calvin. Los organismos heterótrofos obtienen glucosa y galactosa de sus alimentos.
Índice del artículo
- 1 Características
- 2 Tipos de aldosas y sus funciones
- 2.1 La glucosa como fuente principal de energía de los seres vivos
- 2.2 La glucosa y galactosa como componentes en los disacáridos
- 2.3 Glucosa como componente en los polisacáridos
- 2.4 La manosa como componente de las glicoproteínas
- 2.5 La ribosa en el metabolismo
- 2.6 Arabinosa y xilosa como componentes estructurales
- 3 Número de carbonos de las aldosas
- 4 Referencias
En las aldohexosas todos los carbonos son quirales, con excepción del carbono 1, que es el carbono carbonilo del grupo aldehído (C-1), así como el carbono 6, que es un alcohol primario (C-6). Todos los carbonos quirales son alcoholes secundarios.
En todas las aldosas la configuración absoluta del centro quiral más alejado del carbono carbonilo del grupo aldehído puede ser la del D-gliceraldehído o del L-gliceraldehído. Esto determina si la aldosa es un enantiómero D o L.
De manera general, las aldosas con n-carbonos que tienen 2n-2 estereoisomeros. En la naturaleza, las aldosas con la configuración D son más abundantes que las aldosas con configuración L.
La función aldehído de las aldosas reacciona con un grupo hidroxilo secundario en una reacción intramolecular para formar un hemiacetal cíclico. La ciclación de la aldosa convierte el carbono carbonilo en un nuevo centro quiral, denominado carbono anomérico. La posición del sustituyente -OH al carbono anomérico determina la configuración D o L.
Las aldosas cuyo átomo de carbono anomérico no ha formado enlaces glicosídicos son denominadas azúcares reductores. Esto se debe a que las aldosas pueden donar electrones, reduciendo a un agente oxidante o aceptor de electrones. Todas las aldosas son azúcares reductores.
La glicólisis es la vía central universal para el catabolismo de la glucosa. Su función es producir energía en forma de ATP. El piruvato, formado en la glicólisis, puede seguir la vía de fermentación láctica (en músculo esquelético) o la de fermentación alcohólica (en levaduras).
El piruvato también puede ser oxidado completamente a dióxido de carbono mediante un proceso conocido como respiración. Este abarca el complejo de la piruvato deshidrogenasa, el ciclo de Krebs y la cadena transportadora de electrones. En comparación con la fermentación, la respiración produce mucho más ATP por mol de glucosa.
La glucosa está presente en disacáridos tales como celobiosa, isomaltosa, lactosa, maltosa y sacarosa.
La hidrólisis de la lactosa, un azúcar presente en la leche, produce D-glucosa y D-galactosa. Ambos azúcares se encuentran unidos covalentemente mediante un enlace del carbono 1 de la galactosa (configuración β, con el carbono 4 de la glucosa). La lactosa es un azúcar reductor porque el carbono anomérico de la glucosa está disponible, Gal(β1 ->4)Glc.
La sacarosa es uno de los productos de la fotosíntesis y es el azúcar más abundante en muchas plantas. La hidrólisis produce D-glucosa y D-fructosa. La sacarosa no es un azúcar reductor.
La glucosa está presente en los polisacáridos que sirven como substancias de reserva de energía, tales como el almidón y el glicógeno de las plantas y en los mamíferos, respectivamente. También está presente en carbohidratos que sirven de sostén estructural, tales como la celulosa y la quitina de las plantas y los invertebrados, respectivamente.
El almidón es el polisacárido de reserva de las plantas. Se encuentra como gránulos insolubles compuestos por dos tipos de polímeros de glucosa: amilosa y amilopectina.
La amilosa es una cadena sin ramificaciones de residuos de D-glucosa unidos mediante enlaces (α1 -> 4). La amilopectina es una cadena de residuos de glucosa con ramificaciones (α1 -> 6).
El glicógeno es el polisacárido de reserva de los animales. El glicógeno se parece a la amilopectina en que tiene una cadena de residuos de glucosa (α1 -> 4) unidos, pero con muchas más ramificaciones (α1 ->6).
La celulosa forma parte de la pared celular de las plantas, especialmente en los tallos y componentes formadores de madera del cuerpo de la planta. De forma similar a la amilosa, la celulosa es una cadena de residuos de glucosa sin ramificaciones. Posee entre 10,000 y 15,000 unidades de D-glucosa, unidas por enlaces β1 -> 4.
La quitina está conformada por unidades de glucosa modificada, tales como es la N-acetil-D-glucosamina. Se encuentran unidas mediante enlaces β1 -> 4.
Las glicoproteínas poseen uno o varios oligosacáridos. Por lo general, las glicoproteínas se encuentran en la superficie de la membrana plasmática. Los oligosacáridos pueden estar unidos a las proteínas a través de los residuos de serina y treonina (O-unidos) o a residuos de asparagina o glutamina (N-unidos).
Por ejemplo, en plantas, animales y eucariotas unicelulares, el precursor de oligosacárido N-unido es añadido en el retículo endoplasmático. Posee los azúcares siguientes: tres glucosas, nueve manosas y dos N-acetilglucosaminas, lo cual se escribe Glc3Man9(GlcNac)2.
En animales y plantas vasculares, la glucosa puede ser oxidada vía pentosa fosfato para producir ribosa 5-fosfato, una pentosa que formará parte de los ácidos nucleicos. Específicamente, la ribosa llega a formar parte del ARN, mientras que la deoxiribosa llega a formar parte del ADN.
La ribosa también forma parte de otras moléculas, tales como el adenosín trifosfato (ATP), el dinucleótido de adenina y nicotinamida (NADH), el dinucleótido de flavina y adenina (FADH2) y el dinucleótido de adenina y nicotinamida fosforilado (NADPH).
El ATP es una molécula cuya función es aportar energía en diferentes procesos de las células. El NADH y FADH2 participan en el catabolismo de la glucosa, específicamente en reacciones redox. El NADPH es uno de los productos de la oxidación de la glucosa en la vía de la pentosa fosfato. Es fuente de poder reductor en las rutas de biosíntesis de las células.
La pared celular de las plantas está compuesta por celulosa y hemicelulosa. Esta última consiste de heteropolisacáridos que poseen ramificaciones cortas compuestas por hexosas, D-glucosa, D-galactosa y D-manosa, y pentosas como D-xilosa y D-arabinosa.
En las plantas, el ciclo de Calvin es una fuente de azúcares fosforilados, tales como la D-fructosa-6-fosfato, que puede ser transformada en D-glucosa-6-fosfato. Este metabolito es convertido mediante varios pasos, catalizados enzimáticamente, en UDP-xilosa y UDP-arabinosa, que sirven para la biosíntesis de hemicelulosa.
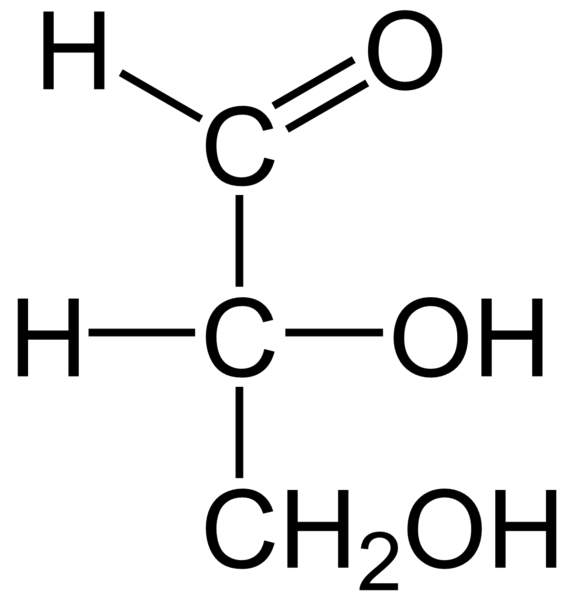
Es una aldosa de tres carbonos, con configuración del D- o L-gliceraldehído. Hay un solo azúcar: el gliceraldehído.
Es una aldosa de cuatro carbonos, con configuración del D- o L-gliceraldehído. Ejemplo: D-eritrosa, D-treosa.
Es una aldosa de cinco carbonos, con configuración del D- o L-gliceraldehído. Ejemplos: D-ribosa, D-arabinosa, D-xilosa.
Es una aldosa de seis carbonos, con configuración del D- o L-gliceraldehído. Ejemplos: D-glucosa, D-manosa, D-galactosa.
- Cui, S. W. 2005. Food carbohydrates: chemistry, physical properties, and applications. CRC Press, Boca Raton.
- Heldt, H. W. 2005. Plant biochemistry. Elsevier, Amsterdam.
- Liptak, A., Szurmai, Z., Fügedi, P., Harangi, J. 1991. CRC handbook of oligosaccharides: volume III: higher oligosaccharides. CRC Press, Boca Raton.
- Lodish, H., et al. 2016. Molecular cell biology. W. H. Freeman, Nueva York.
- Nelson, D. L., Cox, M. M. 2017. Lehninger principles of biochemistry. W. H. Freeman, Nueva York.
- Stick, R. V., Williams, S. J. 2009. Carbohydrates: the essential molecules of life. Elsevier,
- Voet, D., Voet, J. G., Pratt, C. W. 2008. Fundamentals of biochemistry – life at the molecular level. Wiley, Hoboken.