Ciclo del glioxilato: características, reacciones, regulación, funciones
El ciclo del glioxilato es una ruta metabólica presente en las plantas, en algunos microorganismos y en los animales invertebrados (ausente en todos los vertebrados), a través de la cual dichos organismos pueden convertir las grasas en hidratos de carbono (azúcares).
Esta ruta fue descubierta en 1957, mientras Kornberg, Krebs y Beevers trataban de dilucidar cómo bacterias como Escherichia coli podían crecer en presencia de acetato como única fuente de carbono, y cómo las plántulas en germinación del tártago (Ricinus communis) podían convertir las grasas en carbohidratos.
Los estudios de estos tres investigadores llevaron al descubrimiento de dos enzimas conocidas como la isocitrato liasa y la malato sintasa, que, junto con las enzimas del ciclo de Krebs, permiten la síntesis de succinato a partir de dos moléculas de acetil-coA.
El succinato así producido es convertido a malato a través del ciclo de ácidos tricarboxílicos, pudiendo posteriormente ser empleado para la producción de glucosa por medio de la gluconeogénesis.
Esta ruta ocurre, en las plantas, en unos orgánulos especiales denominados glioxisomas y es fundamental para la supervivencia de las plántulas durante las primeras etapas de la germinación.
Índice del artículo
La ruta del glioxilato puede ser contemplada como una “modificación” del ciclo de Krebs, con la diferencia de que en la primera no ocurre una descarboxilación oxidativa, sino que pueden formarse ácidos dicarboxílicos de cuatro átomos de carbono a partir de unidades de acetato de dos carbonos.
Esta característica del ciclo del glioxilato se ha descrito como una forma que tienen algunos organismos para evitar (“bypass”) la pérdida de átomos de carbono en forma de dióxido de carbono que identifica al ciclo de Krebs.
En las plantas, el ciclo del glioxilato ocurre en el interior de unos orgánulos citosólicos rodeados por una membrana simple que se conocen como glioxisomas. En otros organismos como las levaduras y las algas, en cambio, esta ruta ocurre en el citosol.
Los glioxisomas son estructuralmente similares a los peroxisomas (algunos autores los consideran “peroxisomas especializados”), otros orgánulos encargados de parte de la β-oxidación de los ácidos grasos y de la eliminación de las especies reactivas de oxígeno en los organismos eucariotas.
En su interior, los ácidos grasos son oxidados para producir acetil-CoA, que posteriormente se condensa en compuestos de cuatro átomos de carbono. Estos compuestos son selectivamente transportados hacia la mitocondria, donde son convertidos a malato o transportados hacia el citosol para entrar a la ruta gluconeogénica (síntesis de glucosa).
Las enzimas compartidas entre la ruta del glioxilato y el ciclo de ácidos tricarboxílicos existen en la mitocondria y el glioxisoma como isoenzimas, lo que significa que ambas rutas funcionan más o menos independientemente una de la otra.
Los glioxisomas no está presentes en los tejidos vegetales de forma permanente. Son especialmente abundantes durante la germinación de las semillas oleaginosas, las cuales tienen poca capacidad fotosintética para producir los hidratos de carbono que necesitan para crecer.
En las plantas completamente desarrolladas, su participación en el metabolismo de las grasas no es tan imprescindible, pues los azúcares se obtienen principalmente por fotosíntesis.
El acetato proveniente de la degradación de los ácidos grasos funciona como combustible rico en energía y como una fuente de fosfoenolpiruvato para la síntesis de glucosa a través de la gluconeogénesis. El proceso se da como sigue:
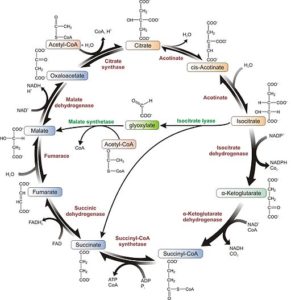
1- La ruta del glioxilato, de forma similar a la del ciclo de Krebs, comienza con la condensación de una molécula de acetil-CoA con otra de oxalacetato para rendir citrato, reacción catalizada por la enzima citrato sintasa.
2- La enzima aconitasa convierte este citrato en isocitrato.
3- El isocitrato es empleado como sustrato de la enzima isocitrato liasa para formar los compuestos succinato y glioxilato.

4- El glioxilato es tomado por la enzima malato sintasa para producir malato a través de su condensación con una segunda molécula de acetil-CoA.
5- El malato es convertido en oxalacetato por la malato deshidrogenasa y dicho compuesto puede servir de precursor para la ruta gluconeogénica o condensarse con otro acetil-CoA para reiniciar el ciclo una vez más.
6- El succinato producido también puede ser convertido a fumarato y este a malato, aportando mayor cantidad de moléculas de oxalacetato para la formación de glucosa. De lo contrario, esta molécula también puede ser exportada hacia la mitocondria para funcionar en el ciclo de Krebs.
El oxalacetato entra a la ruta gluconeogénica para la producción de glucosa gracias a su conversión en fosfoenolpiruvato, que es catalizada por la enzima fosfoenolpiruvato carboxiquinasa.
Puesto que los ciclos del glioxilato y de los ácidos tricarboxílicos comparten numerosos intermediarios entre sí, existe una coordinada regulación entre ambos.
Además, es necesario que existan mecanismos de control, pues la síntesis de glucosa y de otras hexosas a partir del acetil-CoA (proveniente de la degradación de grasas) implica la participación de al menos cuatro rutas:
– La β-oxidación de ácidos grasos que produzca las moléculas de acetil-CoA necesarias tanto para el ciclo del glioxilato como para el ciclo de Krebs y que, en las plantas, tiene lugar en los glioxisomas.
– El ciclo del glioxilato, que también ocurre en los glioxisomas y que, como se dijo, produce intermediarios como el succinato, malato y oxalacetato.
– El ciclo de Krebs, que tiene lugar en la mitocondria y en el cual también se producen los intermediarios succinato, malato y oxalacetato.
– La gluconeogénesis, que ocurre en el citosol y que contempla el uso del oxalacetato convertido en fosfoenolpiruvato para sintetizar glucosa.
El principal punto de control está en la enzima isocitrato deshidrogenasa, cuya regulación implica una modificación covalente por adición o remoción de un grupo fosfato.
Cuando la enzima es fosforilada se inactiva, por lo que el isocitrato se dirige hacia la vía del glioxilato para la producción de glucosa.
Para las plantas el ciclo del glioxilato es fundamental, especialmente durante el proceso de germinación, puesto que la degradación de las grasas almacenadas en las semillas es aprovechada para la síntesis de glucosa en los tejidos poco desarrollados fotosintéticamente hablando.
La glucosa es utilizada como fuente de obtención de energía en forma de ATP o para la formación de carbohidratos más complejos con funciones estructurales, pero algunos de los intermediarios generados durante la ruta del glioxilato también pueden servir propósitos de síntesis de otros componentes celulares.
La principal función del ciclo del glioxilato en los microorganismos es la de proveer una ruta metabólica “alternativa”, de modo que los microorganismos sean capaces de aprovechar otras fuentes de carbono y energía para su crecimiento.
Tal es el caso de la bacteria Escherichia coli, en la cual, cuando los niveles de algunos intermediarios de la glucólisis y del ciclo del ácido cítrico disminuyen (isocitrato, 3-fosfoglicerato, piruvato, fosfoenolpiruvato y oxalacetato), la enzima isocitrato deshidrogenasa (que participa en el ciclo de Krebs) es inhibida y el isocitrato es dirigido hacia la ruta del glioxilato.
Si esta ruta está activa en el momento en que la bacteria crece en un medio rico en acetato, por ejemplo, este metabolito puede ser empleado para sintetizar ácidos carboxílicos de cuatro átomos de carbono que, posteriormente, pueden derivar en la formación de carbohidratos de energéticos.
Para otros organismos como los hongos, por ejemplo, se ha demostrado que la patogenicidad depende en gran medida de la presencia de un ciclo del glioxilato activo, aparentemente por razones metabólicas.
- Dey, P., & Harborne, J. (1977). Plant Biochemistry. San Diego, California: Academic Press.
- Ensign, S. A. (2006). Revisiting the glyoxylate cycle: alternate pathways for microbial acetate assimilation. Molecular Microbiology, 61(2), 274–276.
- Garrett, R., & Grisham, C. (2010). Biochemistry (4th ed.). Boston, USA: Brooks/Cole. CENGAGE Learning.
- Lorenz, M. C., & Fink, G. R. (2001). The glyoxylate cycle is required for fungal virulence. Nature, 412, 83–86.
- Mathews, C., van Holde, K., & Ahern, K. (2000). Biochemistry (3th ed.). San Francisco, California: Pearson.
- Rawn, J. D. (1998). Biochemistry. Burlington, Massachusetts: Neil Patterson Publishers.
- Vallarino, J. G., & Osorio, S. (2019). Organic Acids. In Postharvest Physiology and Biochemistry of Fruits and Vegetables (pp. 207–224). Elsevier Inc.