¿Qué es la anisogamia o heterogamia?
La anisogamia, o heterogamia, es la unión de dos gametos, denominados anisogametos, que difieren en tamaño y estructura. Es el término contrario a la isogamia, que es la unión de dos gametos similares, denominados isogametos.
Dependiendo del tipo de organismo, los anisogametos tienen diferentes características. Por ejemplo, ambos gametos pueden ser flagelados, tal como en algunas algas, o ameboides, tal como ocurre en algunos protozoarios.
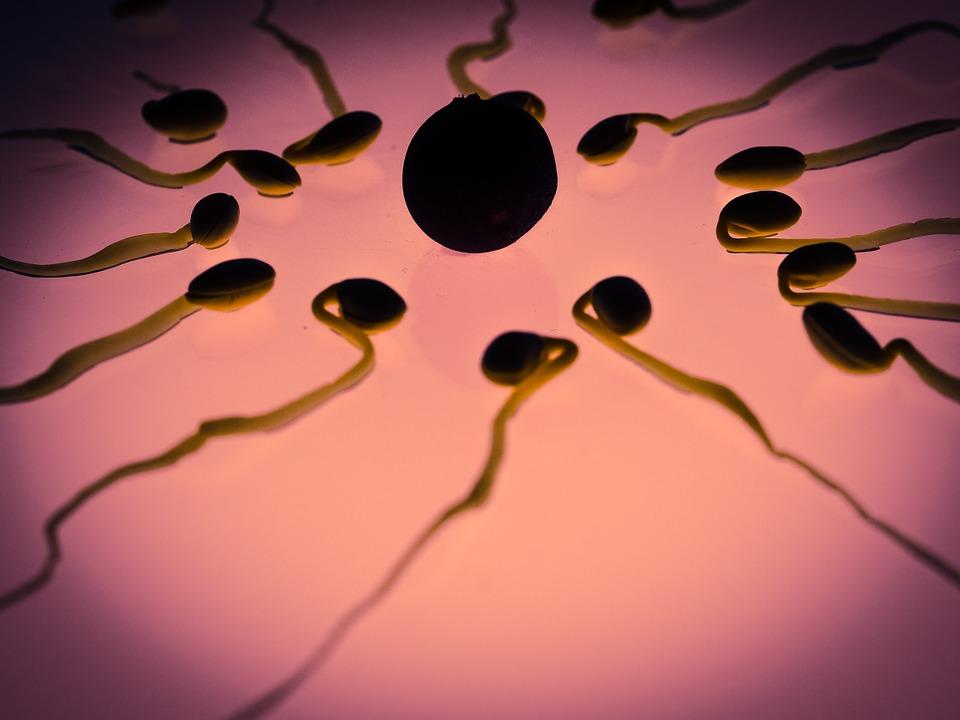
En la oogamia, que es la variante de la anisogamia de los organismos más complejos, un gameto pequeño y móvil, frecuentemente denominado espermatozoide, o polen, fertiliza a un gameto mucho mayor e inmóvil, llamado óvulo.
Índice del artículo
- 1 Aparición de la anisogamia
- 2 Competencia espermática y selección sexual
- 3 Anisogamia y estrategias reproductivas
- 4 Refinamientos reproductivos
- 5 Controversias
- 6 Referencias
La evolución de los organismos multicelulares diploides creó las condiciones necesarias para el desarrollo de la anisogamia, la cual se cree que fue precedida evolutivamente por la isogamia. Los gametos sirven únicamente para la reproducción sexual. La anisogamia permitió especializaciones relacionadas con este tipo de reproducción.
La progresión de la anisogamia hacia su forma más extrema, es decir la oogamia, llevó a la diferenciación de los sexos masculino y femenino, con todo lo que ello implica en términos de dimorfismo, selección y roles sexuales.
A medida que los organismos pluricelulares se fueron haciendo de mayor tamaño, con frecuencia se tornaron escasos. En las plantas y numerosos grupos de animales marinos prevalecieron hábitos sedentarios. Ello creó dificultades para el encuentro de los gametos masculinos y femeninos.
Los machos se especializaron en producir números muy elevados de gametos pequeños (microgametos) capacitados para localizar y fertilizar a los gametos femeninos. Las hembras se especializaron en producir un número reducido de gametos grandes e inmóviles (macrogametos), provistos de materia nutritiva para el desarrollo del embrión.
El exceso de gametos masculinos con respecto a los femeninos inherente a la anisogamia genera una fuerte competencia entre los espermatozoides, o entre el polen, que favorece la selección, tanto entre los gametos masculinos como en los individuos que los producen, de aquellos atributos que favorecen la fertilización.
La selección sexual es el proceso que favorece la evolución de atributos ventajosos para el apareamiento y producción de descendencia por machos y hembras. La selección sexual es responsable de la existencia de las características que distinguen a los sexos. En general, a mayor dimorfismo entre sexos, mayor selección sexual.
En los machos, la selección sexual determina que los gametos masculinos tengan características que incrementan su probabilidad de fertilizar, o que aparezcan atributos anatómicos y conductuales que favorecen el acceso a las hembras al incrementar su capacidad de cortejarlas exitosamente, o de luchar contra otros machos por ellas.
En las hembras, la selección sexual determina la evolución de atributos que les ayudan a escoger a los machos que les permitirán producir descendencia de buena calidad genética, o que posean territorios o aporten recursos alimenticios que favorezcan la crianza.
En muchas especies animales, al buscar pareja, las hembras son selectivas, mientras que los machos no. Esto se atribuye principalmente a que las hembras producen un número limitado de óvulos que deben invertir bien. En contraposición, los machos producen cantidades prácticamente ilimitadas de espermatozoides.
En las especies con cuidado parental, el asunto no es solo una cuestión de óvulos “caros” y espermatozoides “baratos”: las hembras también suelen invertir más en las crías que los machos. La lactancia de los mamíferos, realizada exclusivamente por las hembras, es un ejemplo de ello. Las hembras también arriesgan sus vidas por sus crías.
Si una hembra se aparea con un macho portador de genes defectuosos, y sus hijos no alcanzan la madurez por ello, pierde su esfuerzo reproductivo. En cambio, los machos pueden aparearse con gran número de hembras, pasando sus genes a las generaciones futuras, independientemente del fracaso de algunos de sus descendientes.
Si los machos invierten poco en gametos y en la crianza de sus descendientes, pueden emplear la energía ahorrada para competir con otros machos, e intentar aparearse con el mayor número posible de hembras, maximizando así su capacidad reproductiva. Ello explica la promiscuidad sexual masculina en muchas especies.
Los machos de muchas especies de roedores producen “tapones copulatorios”. El esperma de dichos machos se solidifica en el interior del tracto reproductivo de las hembras impidiendo la copulación a otros machos. Como contramedida, en algunas especies, los machos son capaces de perforar los tapones dejados por otros machos.
En especies en las cuales es común que el esperma de varios machos compita para fecundar los óvulos de una misma hembra, los machos suelen tener testículos y glándulas accesorias mayores, con lo cual producen esperma más concentrado y abundante.
Las hembras han evolucionado mecanismos sofisticados que facilitan o impiden la fertilización por el esperma de diferentes machos. Para ello se valen de contracciones musculares, corrientes ciliares y diversos comportamientos. Las gallinas, por ejemplo, pueden expulsar voluntariamente el esperma de gallos de bajo rango jerárquico.
Charles Darwin consideraba que la existencia de los gametos era uno de los aspectos más enigmáticos de los seres vivos. Un siglo y medio después, la razón de ser de los gametos sigue siendo objeto de debate.
La isogamia es común en los organismos unicelulares. Por el contrario, la anisogamia prevalece en plantas y animales multicelulares. Se ha argumentado que el dimorfismo en tamaño a nivel de gametos permite incrementar el volumen y la complejidad corporal.
Los modelos más aceptados para explicar la anisogamia invocan selección disruptiva: los gametos pequeños se favorecerían porque pueden producirse en gran cantidad; los gametos grandes se favorecerían porque permiten un tamaño mayor de cigoto, lo cual incrementaría la probabilidad de supervivencia del embrión.
Recientemente, algunos autores han cuestionado que la anisogamia sea un factor importante en la evolución de roles sexuales. Según ellos, dichos roles pudieran haber surgido al azar, o como resultado de diferencias en las historias vitales de machos y hembras.
Sin embargo, el consenso ampliamente mayoritario actual es que el azar no puede haber causado diferencias consistentes entre sexos, y que las diferencias en las historias vitales de machos y hembras son el resultado de selección en última instancia determinada por la anisogamia.
- Bell, G. 1978. The evolution of anisogamy. Journal of Theoretical Biology, 73, 247–270.
- Blute, M. 2013. The evolution of anisogamy: more questions than answers. Biological Theory, 7, 3–9.
- Da Silva, J. 2018. The evolution of sexes: a specific test of the disruptive selection theory. Ecology and Evolution, 8, 207–219.
- Kodric-Brown, A., Brown, J. H. 1987. Anisogamy, sexual selection, and the evolution and maintenance of sex. Evolutionary Ecology, 1, 95–105.
- Lehtonen, J., Parker, G. A. 2014. Gamete competition, gamete limitation, and the evolution of the two sexes. Molecular Human Reproduction, 20, 1161–1168.
- Randerson, J. P., Hurst, L. D. 2002. A comparative test of a theory for the evolution of anisogamy. Proceedings: Biological Sciences, 268, 879–884.
- Schärer, L., Rowe, L., Arnqvist, G. 2012. Anisogamy, chance and the evolution of sex roles. Trends in Ecology and Evolution, 2012, 1–5.
- Togashi, T, Cox, P. A., eds. 2011. The evolution of anisogamy – a fundamental phenomenon underlying sexual selection. Cambridge University Press, Cambridge.
- Wedell, N., Gage, M. J. G., Parker, G. A. 2002. Sperm competition, male prudence and sperm-limited females. Trends in Ecology and Evolution, 17, 313–320.