Fosfatidilinositol: estructura, formación, funciones
El fosfatidilinositol es un fosfolípido de la familia de los glicerofosfolípidos o fosfoglicéridos, que están presentes en las membranas biológicas. Constituye cerca del 10% del contenido total de fosfolípidos en una célula promedio.
Es común en la cara interna de la membrana plasmática de muchos eucariotas y procariotas. En los mamíferos y otros vertebrados, enriquece especialmente las membranas de las células cerebrales; y fue en estos tejidos donde fue observado por primera vez por Folch y Wooley en 1942.
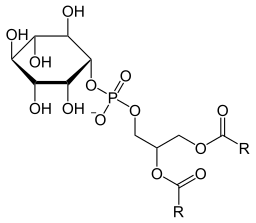
Su estructura química, así como la de algunos de sus derivados fosforilados, fue determinada por el grupo de investigación de Ballou entre los años 1959 y 1961.
Tiene importantes funciones estructurales, relacionadas con su abundancia en las membranas, pero además es una importante fuente de segundos mensajeros que tienen importantes implicaciones en los procesos de señalización celulares que son disparados por numerosos y diversos estímulos especiales.
Su forma glicosilada participa en la modificación covalente de proteínas que les permite unirse a las membranas a través de unas estructuras lipídicas llamadas “anclas” GPI (glicosilfosfatidilinositol).
Índice del artículo
Al igual que la mayor parte de los lípidos de membrana, el fosfatidilinositol es una molécula anfipática, es decir, es una molécula con un extremo polar hidrofílico y otro extremo apolar de naturaleza hidrofóbica.
Su estructura general se basa en un esqueleto de 1,2-diacil glicerol 3-fosfato, donde las dos cadenas de ácidos grasos esterificadas en los carbonos de las posiciones 1 y 2 representan las colas apolares, y el grupo fosfato, unido al grupo “cabeza”, representa la región polar.
Una molécula de inositol unida por medio de un enlace fosfodiéster al grupo fosfato en el carbono de la posición 3 de la molécula de glicerol representa el grupo “cabeza” de este fosfolípido.
El inositol es un derivado del ciclohexano que posee todos sus átomos de carbono (6) unidos cada uno a un grupo hidroxilo. Puede provenir de los alimentos consumidos en la dieta, de la ruta de síntesis de novo o de su propio reciclaje. Las células cerebrales, así como otros tejidos en menor proporción, lo producen a partir de la glucosa 6-fosfato.
La estructura de muchos de los derivados del fosfatidilinositol no son más que una molécula de fosfatidilinositol a la cual se han añadido grupos fosfato en alguno de los grupos hidroxilo de la porción inositol.
Las cadenas hidrocarbonadas de las colas apolares pueden tener longitudes variables desde 16 hasta más o menos 24 átomos de carbono, dependiendo del organismo de que se trate.
Estas cadenas pueden ser saturadas (enlaces sencillos carbono – carbono) o insaturadas (enlaces dobles carbono – carbono; monoinsaturadas o poliinsaturadas) y, al igual que otros fosfolípidos derivados del ácido fosfatídico, la del ácido graso en la posición C2 del glicerol 3-fosfato suele ser insaturado.
Normalmente estos lípidos poseen cadenas de ácidos grasos correspondientes a ácido esteárico y ácido araquidónico, de 18 y 20 átomos de carbono, uno saturado y el otro insaturado, respectivamente.
El fosfatidilinositol, como otros fosfolípidos, es formado a partir de ácido fosfatídico, un fosfolípido sencillo cuya estructura se caracteriza por dos colas apolares y una cabeza polar compuesta únicamente por el grupo fosfato unido al carbono en posición 3 del glicerol.
Para la formación de novo de fosfatidilinositol, el ácido fosfatídico reacciona con CTP (trifosfato de citidina), una molécula de alta energía análoga al ATP, y forma CDP-diacilglicerol, que es un precursor común en la ruta del fosfatidilinositol y sus derivados, del fosfatidilglicerol y del difosfatidilglicerol o la cardiolipina.
La reacción en cuestión es catalizada por la enzima CDP-diacilglicerol sintasa, que tiene una ubicación subcelular dual que involucra a la fracción microsomal y a la membrana mitocondrial interna.
El fosfatidilinositol surge, posteriormente, a partir de una reacción de condensación entre una molécula de inositol y una de CDP-diacilglicerol resultante del paso anterior.
Este paso es catalizado por una fosfatidilinositol sintasa (CDP-diacilglicerol:mioinositol 3-fosfatidil transferasa), una enzima asociada con la membrana del retículo endoplásmico de las células de mamífero.
La reacción que representa el paso limitante de este proceso es en realidad la de la formación del inositol a partir de glucosa 6-fosfato, que debe ocurrir “aguas arriba” de la ruta biosintética.
Los derivados fosforilados del fosfatidilinositol son producidos por un grupo de enzimas denominado fosfatidilinositol quinasas, que se encargan de unir grupos fosfatos a los grupos hidroxilos de la porción inositol del lípido precursor.
Al igual que la fosfatidilserina y el fosfatidilglicerol, el fosfatidilinositol cumple múltiples funciones. Tiene importantes implicaciones estructurales, pues forma parte de las bicapas lipídicas que conforman las diferentes y multifuncionales membranas biológicas.
Muchas proteínas se “unen” a las membranas celulares a través de lo que se denomina “anclas GPI”, que no son más que derivados glicosilados del fosfatidilinositol que le proporcionan a las proteínas un “ancla” hidrofóbica que las sostiene en la superficie de la membrana.
Algunas proteínas del citoesqueleto se unen a los derivados fosforilados del fosfatidilinositol, y este tipo de lípido sirve también como núcleo para la formación de complejos proteicos implicados en la exocitosis.
Sus derivados, por ejemplo, son segundos mensajeros en muchos procesos de señalización relacionados con hormonas en los mamíferos.
Dos de los mensajeros secundarios más importantes que derivan de lo que se ha llamado “el sistema hormona-sensible del fosfatidilinositol” son el inositol 1,4,5-trifosfato (IP3 o inositol trifosfato) y el diacilglicerol, que cumplen diversas funciones “aguas abajo” en la cascada.
El IP3 está implicado en la cascada de señalización de hormonas que emplean los sistemas de segundos mensajeros como, por ejemplo, la adrenalina.
El inositol es un mensajero soluble que ejerce sus funciones en el citosol, mientras que el diacilglicerol es liposoluble y permanece unido en la membrana, donde también ejerce funciones como mensajero.
De la misma forma, en plantas se ha determinado que los derivados fosforilados del fosfatidilinositol también tienen importantes funciones en las cascadas de señalización celulares.
- Antonsson, B. (1997). Phosphatidylinositol synthase from mammalian tissues. Biochimica et Biophysica Acta.
- Luckey, M. (2008). Membrane structural biology: with biochemical and biophysical foundations. Cambridge University Press.
- Murray, R., Bender, D., Botham, K., Kennelly, P., Rodwell, V., & Weil, P. (2009). Harper’s Illustrated Biochemistry (28th ed.). McGraw-Hill Medical.
- Nelson, D. L., & Cox, M. M. (2009). Lehninger Principios de Bioquímica. Ediciones Omega (5th ed.).
- Vance, J. E., & Vance, D. E. (2008). Biochemistry of lipids, lipoproteins and membranes. In New Comprehensive Biochemistry Vol. 36 (4th ed.). Elsevier.