Gliceraldehído 3-fosfato (G3P): estructura, funciones
El gliceraldehído 3-fosfato (GAP) es un metabolito de la glicólisis (su nombre procede del griego; glicos = dulce o azúcar; lisis= ruptura), la cual es una vía metabólica que convierte la molécula de glucosa en dos moléculas de piruvato para producir energía en la forma de adenosín trifosfato (ATP).
En las células, el gliceraldehído 3-fosfato conecta la glicólisis con la gluconeogénesis y la vía de la pentosa fosfato. En los organismos fotosintéticos, el gliceraldehído 3-fosfato, proveniente de la fijación del dióxido de carbono, es utilizado para la biosíntesis de azúcares. En el hígado, el metabolismo de la fructosa produce GAP, el cual es incorporado a la glicólisis.
Índice del artículo
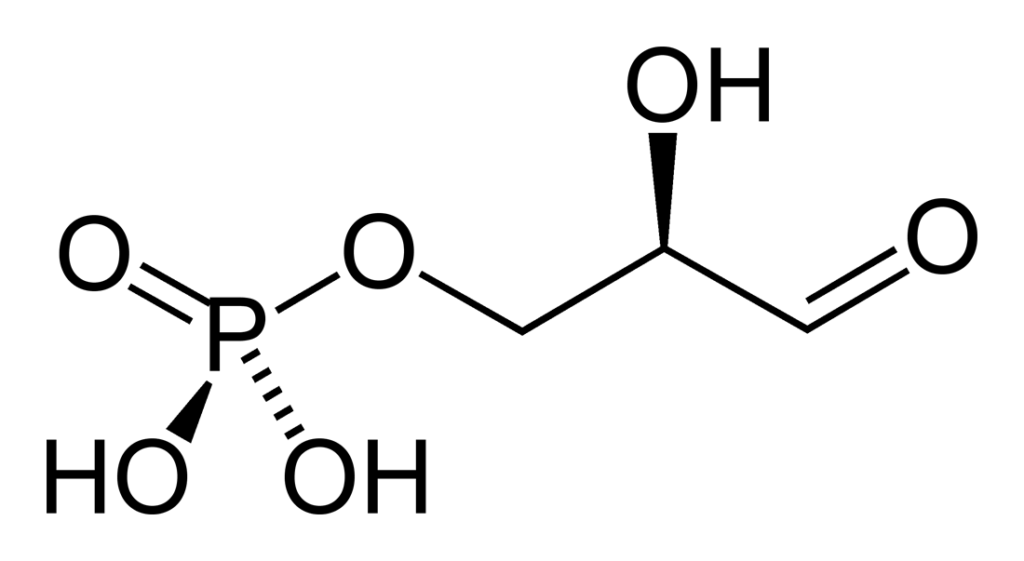
El gliceraldehído 3-fosfato es un azúcar fosforilado que posee tres carbonos. Su fórmula empírica es C3H7O6P. El grupo aldehído (-CHO) es el carbono 1 (C-1), el grupo hidroximetileno (-CHOH) es el carbono 2 (C-2) y el grupo hidroximetil (-CH2OH) es el carbono 3 (C3). Este último forma un enlace con el grupo fosfato (enlace fosfoéster).
La configuración del gliceraldehído 3-fosfato en el C-2 quiral es D. Por convención, respecto al carbono quiral, en la proyección de Fischer el grupo aldehído es representado hacia arriba, el grupo hidroximetil-fosfato hacia abajo, el grupo hidroxilo hacia la derecha y el átomo de hidrógeno hacia la izquierda.
El gliceraldehído 3-fosfato tiene una masa molecular 170,06 g/mol. El cambio de energía libre de Gibbs estándar (ΔGº) para cualquier reacción debe ser calculada sumando la variación de la energía libre de los productos, y restándole la sumatoria de la variación de la energía libre de los reactantes.
De esta manera, se determina la variación energía libre (ΔGº) de formación del gliceraldehído 3-fosfato, que es de -1,285 KJ × mol-1. Por convención, en el estado estándar 25 ºC y 1 atm, la energía libre de los elementos puros es cero.
La glicólisis está presente en todas las células. Se divide en dos fases: 1) etapa de inversión de energía y síntesis de metabolitos con alto potencial de transferencia de grupo fosfato, como es el gliceraldehído 3-fosfato (GAP); 2) etapa de síntesis de ATP a partir de moléculas con alto potencial de transferencia de grupo fosfato.
El gliceraldehído 3-fosfato y la dihidroxiacetona fosfato se forman a partir de la fructosa 1,6-bifosfato, una reacción catalizada por la enzima aldolasa. El gliceraldehído 3-fosfato es convertido en 1,3-bifosfoglicerato(1,3BPG), mediante una reacción catalizada por la enzima GAP deshidrogenasa.
La GAP deshidrogenasa cataliza la oxidación del átomo de carbono del aldehído y transfiere un grupo fosfato. Así, se forma un anhídrido mixto (el 1,3BPG) en el que el grupo acilo, y el átomo de fósforo quedan propenso a la reacción de ataque nucleofílico.
Seguidamente, en una reacción catalizada por la 3-fosfoglicerato quinasa, el 1,3BPG transfiere el grupo fosfato desde el carbono 1 al ADP, formándose ATP.
Debido a que las reacciones catalizadas por la aldolasa, la GAP deshidrogenasa y la 3-fosfoglicerato quinasa están en el equilibrio (ΔGº ~ 0), son reversibles, siendo por ende parte de la vía de la gluconeogénesis (o de la síntesis nueva de glucosa).
En la vía de la pentosa fosfato, el gliceraldehído 3-fosfato (GAP) y la fructosa 6-fosfato (F6P) se forman mediante reacciones de corte y formación de enlaces C-C, a partir de las pentosas, xilulosa 5-fosfato y ribosa 5-fosfato.
El gliceraldehído 3-fosfato puede seguir la vía de la gluconeogenesis y formar glucosa 6-fosfato, que continúa la vía de la pentosa fosfato. La glucosa puede ser oxidada por completo produciendo seis moléculas de CO2 a través de la etapa oxidativa de la vía pentosa fosfato.
En el ciclo de Calvin, el CO2 es fijado como 3-fosfoglicerato, en una reacción catalizada por la ribulosa bifosfato carboxilasa. Luego, el 3-fosfoglicerato es reducido por el NADH mediante la acción de una enzima denominada GAP deshidrogenasa.
Se necesita 2 moléculas de GAP para la biosíntesis de una hexosa, tal como la glucosa, la cual sirve para la biosíntesis de almidón o celulosa en las plantas.
La enzima fructoquinasa cataliza la fosforilación de la fructosa por el ATP en C-1, formando fructosa 1-fosfato. La aldolasa A, que se encuentra en el músculo, es específica para la fructosa 1,6-bifosfato como sustrato. La aldolasa B se encuentra en el hígado y es específica para la fructosa 1-fosfato como sustrato.
La aldolasa B cataliza la ruptura aldólica de la fructosa 1-fosfato y produce dihidroxiacetona fosfato y gliceraldehído. La gliceraldehído quinasa cataliza la fosforilación del gliceraldehído mediante ATP, formando un intermediario glicolítico, el gliceraldehído 3-fosfato (GAP).
En una vía diferente, el gliceraldehído es transformado en glicerol por una alcohol deshidrogenasa que usa NADH como sustrato donador de electrones. Luego, la glicerol quinasa fosforila el glicerol mediante ATP, formando glicerol fosfato. Este último metabolito es reoxidado, formando dihidroxiacetona fosfato (DHAP) y NADH.
La DHAP es convertida en GAP por la enzima triosa fosfato isomerasa. De esta manera, la fructosa es convertida en metabolitos de la glicólisis. Sin embargo, la fructosa suministrada de forma intravenosa puede ocasionar daños graves, los cuales consisten de la disminución drástica de fosfato y ATP intracelular. Inclusive se produce acidosis láctica.
Los daños de la fructosa se deben a que no tiene los puntos de regulación que normalmente tiene el catabolismo de la glucosa. En primer lugar, la fructosa entra a los músculos a través de GLUT5, que es independiente de insulina.
En segundo lugar, la fructosa es convertida directamente en GAP y de esta manera no pasa por la regulación de la enzima fosfofructo quinasa (PFK) al comienzo de la glicólisis.
La glicólisis es la vía universal para el catabolismo de la glucosa. Sin embargo, algunas bacterias utilizan alternativamente la vía Entner-Doudoroff. Esta vía implica seis pasos catalizados por enzimas, en los cuales la glucosa es transformada en GAP y piruvato, que son dos productos finales de esta vía.
El GAP y el piruvato son transformados en etanol mediante reacciones de fermentación alcohólica.
- Berg, J. M., Tymoczco, J. L., Stryer, L. 2015. Biochemistry. A short course. W. H. Freeman, Nueva York.
- Miesfeld, R. L., McEvoy, M. M. 2017. Biochemistry. W. W. Norton, Nueva York.
- Nelson, D. L., Cox, M. M. 2017. Lehninger principles of biochemistry. W. H. Freeman, Nueva York.
- Salway J. G. 2004. Metabolism at a glance. Blackwell, Malden.
- Voet, D., Voet, J. G., Pratt, C. W. 2008. Fundamentals of biochemistry: life at the molecular level. Wiley, Hoboken.