Glicerol 3-fosfato: estructura, características, funciones
El glicerol 3-fosfato es una molécula de glicerol que posee un enlace éster con un grupo fosfato, teniendo numerosas funciones en el metabolismo y formando parte de las biomembranas. Esta molécula sirve como metabolito para la gluconeogénesis, la biosíntesis de triacilgliceroles y la biosíntesis de segundos mensajeros como el diacilglicerol (DAG).
Otras funciones del glicerol 3-fosfato son la biosíntesis de glicerofosfolípidos, tales como la cardiolipina, plasmalógenos, y alquilacilglicerofosfolípidos. Además, participa en una lanzadera que permite regenerar NAD+ en el citosol.
Índice del artículo
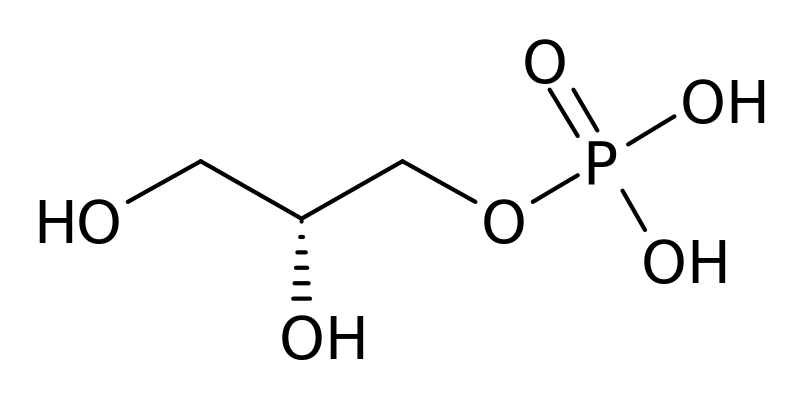
La fórmula empírica del glicerol 3-fosfato es C3H9O6P y posee tres átomos de carbono. Los átomos de carbono 1 y 3 (C-1 y C-3) forman grupos hidroximetil (-CH2OH), mientras que el átomo de carbono 2 (C-2) forma un grupo hidroximetileno (-CHOH). El átomo de oxígeno del grupo hidroximetil, de C-3, forma un enlace éster con un grupo fosfato.
Hay sinónimos del glicerol 3-fosfato, tales como 1,2,3-propanetriol, 1-(dihidrógeno fosfato) y 2,3-dihidroxipropil dihidrógeno fosfato, 3-fosfoglicerol. Su peso molecular es 172,07 g/mol.
El cambio de energía libre de Gibbs estándar (ΔGº) de la hidrólisis del grupo fosfato del glicerol 3-fosfato es -9,2 KJ/mol.
Este metabolito es convertido en un intermediario de la glicólisis. Cuando la carga energética celular es alta, el flujo a través de la glicólisis se ve disminuido y la dihidroxiacetona fosfato (DHAP) sirve como material de partida para las vías de biosíntesis.
El glicerol sirve como metabolito para las vías anabólicas. Para ello, debe ser convertido en un intermediario glicolítico mediante dos pasos que necesitan las enzimas glicerol quinasa y la glicerol fosfato deshidrogenasa para formar el intermediario dihidroxiacetona-fosfato (DHAP).
La enzima glicerol quinasa cataliza la transferencia de un grupo fosfato desde el ATP (adenosín trifosfato) al glicerol, formándose glicerol 3-fosfato y ADP (adenosín difosfato). A continuación, la glicerol 3-fosfato deshidrogenasa cataliza una reacción de oxido-reducción, en la cual el C-2 del glicerol 3-fosfato es oxidado, perdiendo dos electrones.
Los electrones del glicerol 3-fosfato (reducido) son cedidos al NAD+ (oxidado), formando DHAP (oxidado) y NADH (reducido). El DHAP es un metabolito intermediario de la glicólisis que provee esqueletos carbonados para las vías anabólicas, tales como la de la biosíntesis de glicógeno y nucleótidos.
La glucosa 6-fosfato formada mediante gluconeogénesis puede continuar hacia la biosíntesis de glicógeno o hacia la vía de la pentosa fosfato. Durante la biosíntesis de glucógeno en el hígado, la glucosa 6-fosfato es convertida en glucosa 1-fosfato. Durante la vía de la pentosa fosfato, la glucosa 6-fosfato es convertida en ribosa 5-fosfato.
Los triacilgliceroles son lípidos neutros (no poseen carga) que poseen ésteres de ácidos grasos covalentemente unidos al glicerol. Los triacilgliceroles son sintetizados a partir de ésteres de acilgrasos-CoA y glicerol 3-fosfato o DHAP.
La gliceroneogénesis es la biosíntesis nueva del glicerol a partir del oxalacetato, usando enzimas de la gluconeogénesis. La piruvato carboxilasa transforma el piruvato en oxaloacetato, y la fosfoenolpiruvato carboxiquinasa (PEPCK) transforma el oxaloacetato en fosfoenolpiruvato, un intermediario glicolítico.
El fosfoenolpiruvato continúa la vía de la gluconeogénsis hacia la biosíntesis de DHAP, el cual es convertido en glicerol mediante la glicerol 3-fosfato deshidrogenasa y una fosfatasa que hidroliza el grupo fosfato. El glicerol así formado es utilizado para la biosíntesis de triacilgliceroles.
Durante períodos de inanición, el 30% de los ácidos grasos que entran al hígado son reesterificados a triacilgliceroles y exportados como lipoproteínas de muy baja densidad (VLDL).
Los adipocitos, aunque no llevan a cabo gluconeogénesis, poseen la enzima fosfoenolpiruvato carboxiquinasa (PEPCK), la cual participa en la glicerogénesis necesaria para la biosíntesis de triacilglicerol.
Los glicerofosfolípidos son triésteres de glicerol 3-fosfato, en los cuales el fosfato es la cabeza polar. Los C-1 y C-2 forman enlaces éster con ácidos grasos saturados, tal como el palmitato o el esterato, y un ácido graso monoinsaturado, tal como el oleato. Esta descripción corresponde al fosfatidato, que es el glicerofosfolípido más simple.
En las membranas celulares de los eucariotas, el fosfatidato sirve como precursor de los glicerofosfolípidos más comunes, que son la fosfatidilcolina, la fosfatidilserina, la fosfatidiletanolamina y el fosfatidilinositol.
La distribución de los lípidos (glicerofosfolípidos, esfingofosfolípidos, esfingoglicolípidos, colesterol) en las membranas celulares no es uniforme. Por ejemplo, la monocapa interna de la membrana de los eritrocitos es rica en glicerofosfolípidos, mientras que la monocapa externa es rica en esfingolípidos.
Los glicerofosfolípidos son importantes porque participan en el señalamiento celular. Mediante la acción de enzimas fosfolipasas, tales como la fosfolipasa C que rompe el enlace éster a nivel del C-3 del fosfatidilinositol-4,5-bifosfato (PPI2), se producen las moléculas de señalamiento inositol 1,4,5 -trifosfato y diacilglicerol (DAG).
Frecuentemente, los venenos de serpientes contienen enzimas fosfolipasa A2, las cuales rompen los glicerofosfolípidos. Esto produce daño a los tejidos por ruptura de las membranas. Los ácidos grasos liberados actúan como detergentes.
Las membranas de las células eucariotas contienen otros fosfolípidos como son la cardiolipina, plasmalógenos y alquilacilglicerofosfolípidos.
La cardiolipina es un fosfolípido que fue aislado por primera vez del tejido del corazón. Su biosíntesis requiere dos moléculas de fosfatidilglicerol. Los plasmalógenos contienen cadenas hidrocarbonadas unidas al C-1 del glicerol mediante un enlace éter vinilo. En los mamíferos, el 20% de los glicerofosfolípidos son plasmalógenos.
En los alquilacilglicerofosfolípidos un sustituyente alquil está unido al C-1 del glicerol mediante unión éter. Estos glicerofosfolípidos son menos abundantes que los plasmalógenos.
El músculo esquelético, el cerebro y el músculo de los insectos voladores usan la lanzadera glicerol 3-fosfato. El glicerol 3-fosfato consiste principalmente de dos isoenzimas: el glicerol 3-fosfato deshidrogenasa y una flavoproteína deshidrogenasa.
La glicerol 3-fosfato deshidrogenasa cataliza la oxidación del NADH citosólico. Este NADH se produce en la glicólisis, en el paso catalizado por la gliceraldehído 3-fosfato deshidrogenasa (GAPDH). La glicerol 3-fosfato deshidrogenasa cataliza la transferencia de dos electrones desde el NADH (reducido) hacia el sustrato dihidroxiacetona-fosfato (oxidado).
Los productos de la catálisis de la glicerol 3-fosfato deshidrogenasa son el NAD+ (oxidado) y el glicerol 3-fosfato (reducido). Este último es oxidado por una flavoproteína deshidrogenasa que se encuentra en la membrana interna de la mitocondria. De esta manera, el DHAP es reciclado.
La flavoproteína deshidrogenasa cede los electrones a la cadena transportadora de electrones. Debido a ello, el NADH del citosol sirve para la biosíntesis de 1,5 moléculas de ATP mediante fosforilación oxidativa en la cadena transportadora de electrones. La regeneración del NAD+ en el citosol permite que la glicósis continúe. La GAPDH usa NAD+ como sustrato.
- Berg, J. M., Tymoczco, J. L., Stryer, L. 2015. Biochemistry: a short course. W. H. Freeman, Nueva York.
- Lodish, H., Berk, A., Zipurski, S. L., Matsudaria, P., Baltimore, D., Darnell, J. 2003. Biología celular y molecular. Editorial Médica Panamericana, Buenos Aires.
- Miesfeld, R. L., McEvoy, M. M. 2017. Biochemistry. W. W. Norton, Nueva York.
- Nelson, D. L., Cox, M. M. 2017. Lehninger principles of biochemistry. W. H. Freeman, Nueva York.
- Voet, D., Voet, J. G., Pratt, C. W. 2008. Fundamentals of biochemistry: life at the molecular level. Wiley, Hoboken.