Síntesis de ácidos grasos: dónde ocurre, enzimas, etapas y reacciones
La síntesis de ácidos grasos es el proceso por el cual se producen los componentes fundamentales de los lípidos más importantes de las células (los ácidos grasos), que participan en muchas funciones celulares muy relevantes.
Los ácidos grasos son moléculas alifáticas, es decir, están compuestos esencialmente de átomos de carbono e hidrógeno unidos entre sí de manera más o menos lineal. Tienen un grupo metilo en uno de sus extremos terminales y un grupo carboxílico ácido en el otro, por el que son denominados “ácidos grasos”.
Los lípidos son moléculas empleadas por distintos sistemas biosintéticos celulares para la formación de otras moléculas más complejas como:
- los fosfolípidos de membrana
- los triglicéridos para el almacenamiento de energía y
- las anclas de algunas moléculas especiales que se encuentran en la superficie de muchos tipos de células (eucariotas y procariotas)
Estos compuestos pueden existir como moléculas lineales (con todos los átomos de carbono saturados con moléculas de hidrógeno), pero también pueden observarse los de cadena lineal y con algunas saturaciones, es decir, con dobles enlaces entre sus átomos de carbono.
Los ácidos grasos saturados también pueden encontrarse con cadenas ramificadas, cuya estructura es un poco más compleja.
Las características moleculares de los ácidos grasos son cruciales para su función, pues de ellas dependen muchas de las propiedades fisicoquímicas de las moléculas que están formadas por estos, especialmente de su punto de fusión, su grado de empaquetamiento y su capacidad de formación de bicapas.
Así, la síntesis de ácidos grasos es un asunto sumamente regulado, pues se trata de una serie de eventos secuenciales críticos para la célula desde muchos puntos de vista.
Índice del artículo
- 1 ¿Dónde ocurre la síntesis de ácidos grasos?
- 2 Enzimas que participan
- 3 Etapas y reacciones
- 4 Regulación
- 5 Referencias
En la mayor parte de los organismos vivos la síntesis de los ácidos grasos se da en el compartimento citosólico, mientras que su degradación ocurre principalmente entre el citosol y la mitocondria.
El proceso depende de la energía contenida en los enlaces del ATP, del poder reductor del NADPH (usualmente derivado de la ruta de las pentosas fosfato), del cofactor biotina, de iones bicarbonato (HCO3-) y de iones de manganeso.
En los animales mamíferos los principales órganos de síntesis de ácidos grasos son el hígado, los riñones, el cerebro, los pulmones, las glándulas mamarias y el tejido adiposo.
El sustrato inmediato de la síntesis de novo de los ácidos grasos es el acetil-CoA y el producto final es una molécula de palmitato.
El acetil-CoA deriva directamente del procesamiento de los intermediarios glucolíticos, razón por la cual una dieta alta en carbohidratos promueve la síntesis de lípidos (lipogénesis) ergo, también de ácidos grasos.
El acetil-CoA es el bloque de síntesis de dos carbonos que se emplea para la formación de los ácidos grasos, puesto que varias de estas moléculas se unen consecutivamente a una molécula de malonil-CoA, formada por la carboxilación de un acetil-CoA.
La primera enzima de la ruta, y una de las más importantes desde el punto de vista de la regulación de la misma, es aquella encargada de la carboxilación del acetil-CoA, conocida como acetil-CoA carboxilasa (ACC), que es un complejo enzimático formado por 4 proteínas y que utiliza biotina como cofactor.
No obstante, y muy a pesar de que existen diferencias estructurales entre las distintas especies, la enzima ácido graso sintasa es la que se encarga de las principales reacciones biosintéticas.
Esta enzima es, en realidad, un complejo enzimático compuesto por monómeros que tienen las 7 actividades enzimáticas distintas, que son necesarias para la elongación del ácido graso en “nacimiento”.
Las 7 actividades de esta enzima se pueden enlistar como sigue:
– ACP: proteína transportadora de grupo acilo
– Acetil-CoA-ACP transacetilasa (AT)
– β-cetoacil-ACP sintasa (KS)
– Malonil-CoA-ACP transferasa (MT)
– β-cetoacil-ACP reductasa (KR)
– β-hidroxiacil-ACP deshidratasa (HD)
– Enoil-ACP reductasa (ER)
En algunos organismos como las bacterias, por ejemplo, el complejo ácido graso sintasa está formado por proteínas independientes que se asocian entre sí, pero que son codificadas por genes distintos (sistema de ácido graso sintasa tipo II).
No obstante, en muchos eucariotas y algunas bacterias la multienzima contiene varias actividades catalíticas que están separadas en dominios funcionales distintos, en uno o más polipéptidos, pero que pueden ser codificadas por el mismo gen (sistema de ácido graso sintasa tipo I).
La mayor parte de los estudios realizados respecto a la síntesis de ácidos grasos involucra los hallazgos realizados en el modelo bacteriano, no obstante, los mecanismos de síntesis de los organismos eucariotas también han sido estudiados con cierta profundidad.
Es importante mencionar que el sistema ácido graso sintasa tipo II se caracteriza porque todos los intermediarios acil graso se unen covalentemente a una proteína acídica de pequeño tamaño conocida como la proteína transportadora de acilo (ACP), que los transporta de una enzima a la siguiente.
En los eucariotas, por el contrario, la actividad ACP es parte de la misma molécula, entendiéndose que la misma enzima tiene un sitio especial para la unión de los intermediarios y su transporte a través de los distintos dominios catalíticos.
La unión entre la proteína o la porción ACP y los grupos acil graso ocurre a través de enlaces tioéster entre estas moléculas y el grupo prostético 4’-fosfopanteteína (ácido pantoténico) de la ACP, que se condensa con el grupo carboxilo del acil graso.
- Inicialmente la enzima acetil-CoA carboxilasa (ACC) se encarga de catalizar el primer paso de “compromiso” en la síntesis de los ácidos grasos que, como se comentó, implica la carboxilación de una molécula de acetil-CoA para formar el intermediario de 3 átomos de carbono conocido como malonil-CoA.
El complejo ácido graso sintasa recibe los grupos acetilo y malonil, que deben “rellenar” correctamente los sitios “tiol” de esta.
Esto tiene lugar inicialmente por la transferencia del acetil-CoA al grupo SH de la cisteína en la enzima β-cetoacil-ACP sintasa, reacción catalizada por la acetil-CoA-ACP transacetilasa.
El grupo malonil es transferido del malonil-CoA al grupo SH de la proteína ACP, evento mediado por la enzima malonil-CoA-ACP transferasa, formando malonil-ACP.
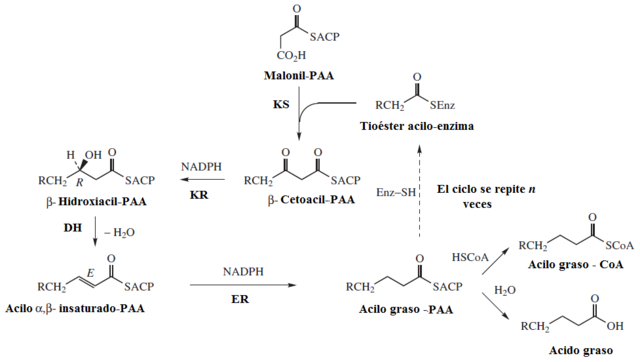
- El paso de iniciación de la elongación del ácido graso en nacimiento consiste en la condensación del malonil-ACP con una molécula de acetil-CoA, reacción dirigida por una enzima con actividad β-cetoacil-ACP sintasa. En esta reacción se forma, entonces, acetoacetil-ACP y se libera una molécula de CO2.
- Las reacciones de elongación ocurren en ciclos donde se adicionan 2 átomos de carbono por vez, en que cada ciclo consiste en una condensación, una reducción, una deshidratación y un segundo evento de reducción:
– Condensación: se condensan los grupos acetilo y malonil para formar el acetoacetil-ACP
– Reducción del grupo carbonilo: el grupo carbonilo del carbono 3 del acetoacetil-ACP es reducido, formándose D-β-hidroxibutiril-ACP, reacción catalizada por la β-cetoacil-ACP-reductasa, que emplea NADPH como donador de electrones.
– Deshidratación: se remueven los hidrógenos entre los carbonos 2 y 3 de la molécula anterior, formando un doble enlace que termina con la producción del trans-∆2-butenoil-ACP. La reacción es catalizada por la β-hidroxiacil-ACP deshidratasa.
– Reducción del doble enlace: el doble enlace del trans-∆2-butenoil-ACP es reducido para formar butiril-ACP por acción de la enoil-ACP reductasa, que también emplea NADPH como agente reductor.
Para continuar con la elongación, una nueva molécula de malonil debe unirse nuevamente a la porción ACP del complejo ácido graso sintasa y comienza con la condensación de esta con el grupo butiril formado en el primer ciclo de síntesis.
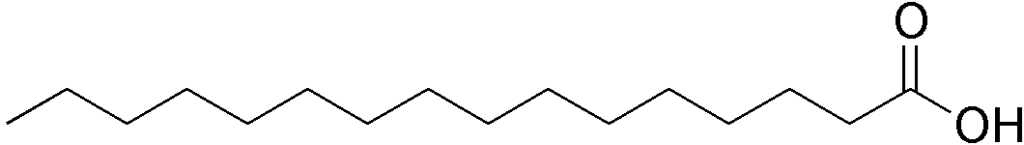
En cada paso de elongación se emplea una nueva molécula de malonil-CoA para crecer la cadena en 2 átomos de carbono y estas reacciones se repiten hasta que se alcanza la longitud adecuada (16 átomos de carbono), luego de lo cual una enzima tioesterasa libera el ácido graso completo por hidratación.
El palmitato puede ser procesado ulteriormente por distintos tipos de enzimas que modifican sus características químicas, es decir, que pueden introducir insaturaciones, prolongar su longitud, etc.
Al igual que muchas rutas biosintéticas o de degradación, la síntesis de ácidos grasos es regulada por distintos factores:
– Depende de la presencia de iones bicarbonato (HCO3-), de vitamina B (biotina) y de acetil-CoA (durante el paso inicial de la ruta, que implica la carboxilación de una molécula de acetil-CoA por medio de un intermediario carboxilado de biotina para formar malonil-CoA).
– Es una ruta que ocurre en respuesta a las características energéticas celulares, pues cuando existe suficiente cantidad de “combustible metabólico”, el exceso es convertido en ácidos grasos que son almacenados para su posterior oxidación en momentos de déficit energético.
En términos de la regulación de la enzima acetil-CoA carboxilasa, que representa el paso limitante de toda la ruta, esta es inhibida por el palmitoil-CoA, el principal producto de la síntesis.
Su activador alostérico, en cambio, es el citrato, que dirige el metabolismo desde la oxidación hacia la síntesis para su almacenamiento.
Cuando las concentraciones mitocondriales de acetil-CoA y de ATP incrementan, el citrato es transportado hacia el citosol, donde es tanto precursor para la síntesis citosólica de acetil-CoA como una señal alostérica de activación para la acetil-CoA carboxilasa.
Esta enzima también puede ser regulada por fosforilación, evento disparado por la acción hormonal del glucagón y la epinefrina.
- McGenity, T., Van Der Meer, J. R., & de Lorenzo, V. (2010). Handbook of hydrocarbon and lipid microbiology (p. 4716). K. N. Timmis (Ed.). Berlin: Springer.
- Murray, R. K., Granner, D. K., Mayes, P. A., & Rodwell, V. W. (2014). Harper’s illustrated biochemistry. Mcgraw-hill.
- Nelson, D. L., & Cox, M. M. (2009). Lehninger principles of biochemistry (pp. 71-85). New York: WH Freeman.
- Numa, S. (1984). Fatty acid metabolism and its regulation. Elsevier.
- Rawn, J. D. (1989). Biochemistry—International edition. North Carolina: Neil Patterson Publishers, 5.