Bomba de calcio: funciones, tipos, estructura y funcionamiento
La bomba de calcio es una estructura de naturaleza proteica que se encarga del transporte de calcio a través de membranas de las células. Esta estructura es dependiente de ATP y se considera una proteína de tipo ATPasa, también llamada Ca2+-ATPasa.
La Ca2+-ATPasa se encuentra en todas las células de los organismos eucariotas y son esenciales para la homeostasis de calcio en la célula. Esta proteína lleva a cabo un transporte activo primario, ya que el movimiento de las moléculas de calcio va en contra de su gradiente de concentración.
Índice del artículo
El Ca2+ cumple papeles importantes en la célula, por lo que su regulación dentro de ellas es fundamental para su correcto funcionamiento. A menudo, actúa como un segundo mensajero.
En los espacios extracelulares la concentración de Ca2+ es aproximadamente 10.000 veces mayor que dentro de las células. Un aumento de la concentración de este ión en el citoplasma celular desencadena diversas respuestas, como contracciones musculares, liberación de neurotransmisores y la degradación de glucógeno.
Existen varias formas de trasladar estos iones desde las células: transporte pasivo (salida inespecífica), canales iónicos (movimiento a favor de su gradiente electroquímico), transporte activo secundario de tipo antiporte (Na/Ca), y el transporte activo primario con la bomba dependiente de ATP.
A diferencia de los otros mecanismos de desplazamiento de Ca2+, la bomba funciona en forma vectorial. Es decir, el ion se mueve en una sola dirección de manera que solo funciona expulsándolos.
La célula es extremadamente sensible a los cambios de concentración del Ca2+. Al presentar una diferencia tan marcada con su concentración extracelular, por ello es tan importante restablecer eficientemente sus niveles citosólicos normales.
Se han descrito tres tipos de Ca2+-ATPasas en las células de animales, según sus ubicaciones en las células; las bombas ubicadas en la membrana plasmática (PMCA), las ubicadas en el retículo endoplasmático y la membrana nuclear (SERCA), y las encontradas en la membrana del aparato de Golgi (SPCA).
Las bombas SPCA también transportan iones Mn2+ que son cofactores de diversas enzimas de la matriz del aparato de Golgi.
Las células de levaduras, de otros organismos eucariontes y las células vegetales presentan otros tipos de Ca2+-ATPasas muy particulares.
En la membrana plasmática encontramos el transporte activo antipórtico Na/Ca, siendo el responsable del desplazamiento de una importante cantidad de Ca2+ en las células en reposo y actividad. En la mayoría de células en estado de reposo, el encargado del transporte de calcio hacia el exterior es la bomba PMCA.
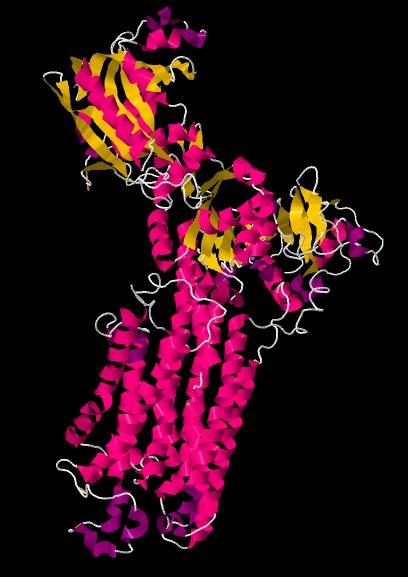
Estas proteínas están conformadas por unos 1.200 aminoácidos, y posee 10 segmentos transmembranales. En el citosol se encuentran 4 unidades principales. La primera unidad contiene el grupo amino-terminal. La segunda tiene características básicas, lo que permite que se una a los fosfolípidos ácidos activadores.
En la tercera unidad se encuentra un ácido aspártico con función catalítica, y “aguas abajo” de este una banda de unión de isotocianato de fluoresceína, en el dominio de unión de ATP.
En la cuarta unidad se encuentra el dominio de unión a la calmodulina, los sitios de reconocimiento de ciertas quinasas (A y C) y las bandas de unión de Ca2+ alostérico.
Las bombas SERCA se encuentran en gran cantidad en el retículo sarcoplásmico de las células musculares y su actividad está relacionada con la contracción y relajación en el ciclo del movimiento muscular. Su función es transportar el Ca2+ desde el citosol de la célula hasta la matriz del retículo.
Estas proteínas consisten en una única cadena polipeptídica con 10 dominios transmembranales. Su estructura es básicamente igual al de las proteínas PMCA, pero difiere en que estas solo poseen tres unidades dentro del citoplasma, encontrándose el sitio activo en la tercera unidad.
El funcionamiento de esta proteína requiere un balance de cargas durante el transporte de los iones. Dos Ca2+ (por ATP hidrolizado) son desplazados desde el citosol hasta la matriz del retículo, en contra de un gradiente de concentración muy alto.
Este transporte ocurre de manera antipórtica, pues al mismo tiempo dos H+ son dirigidos al citosol desde la matriz.
El mecanismo de transporte se divide en dos estados E1 y E2. En el E1 los sitios de unión que presentan una alta afinidad por el Ca2+ están dirigidos hacia el citosol. En el E2 los sitios de unión se dirigen hacia el lumen del retículo presentando una baja afinidad por Ca2+. Los dos iones Ca2+ se unen tras la transferencia.
Durante la unión y transferencia de los Ca2+, ocurren cambios conformacionales, entre ellos la apertura del dominio M de la proteína, que está hacia el citosol. Los iones se unen entonces de manera más sencilla a los dos sitios de unión de dicho dominio.
La unión de los dos iones Ca2+ promueve una serie de cambios estructurales en la proteína. Entre ellos la rotación de ciertos dominios (dominio A) que reorganiza las unidades de la bomba, habilitando la apertura hacia la matriz del retículo para liberar los iones, que se desacoplan gracias a la disminución de la afinidad en los sitios de unión.
Los protones H+ y las moléculas de agua estabilizan el sitio de unión de los Ca2+, ocasionando que el dominio A rote de nuevo a su estado original, cerrando el acceso hacia el retículo endoplasmático.
Este tipo de bombas se encuentran en todas las células eucariotas y se encarga de la expulsión de Ca2+ hacia el espacio extracelular con el fin de mantener estable su concentración dentro de las células.
En esta proteína se transporta un ion de Ca2+ por ATP hidrolizado. El transporte está regulado por los niveles de la proteína calmodulina en el citoplasma.
Al aumentar la concentración de Ca2+ citosólico, aumentan los niveles de calmodulina, que se unen a los iones de calcio. El complejo Ca2+-calmodulina, se ensambla entonces al sitio de unión de la bomba PMCA. Se produce un cambio conformacional en la bomba que permite que se exponga la apertura hacia el espacio extracelular.
Los iones de calcio se liberan, restaurando los niveles normales en el interior de la célula. Consecuentemente el complejo Ca2+-calmodulina se desensambla, regresando la conformación de la bomba a su estado original.
- Brini, M., & Carafoli, E. (2009). Calcium pumps in health and disease. Physiological reviews, 89(4), 1341-1378.
- Carafoli, E., & Brini, M. (2000). Calcium pumps: structural basis for and mechanism of calcium transmembrane transport. Current opinion in chemical biology, 4(2), 152-161.
- Devlin, T. M. (1992). Textbook of biochemistry: with clinical correlations.
- Latorre, R. (Ed.). (1996). Biofísica y fisiología celular. Universidad de Sevilla.
- Lodish, H., Darnell, J. E., Berk, A., Kaiser, C. A., Krieger, M., Scott, M. P., & Matsudaira, P. (2008). Mollecular cell biology. Macmillan.
- Pocock, G., & Richards, C. D. (2005). Fisiología humana: la base de la medicina. Elsevier España.
- Voet, D., & Voet, J. G. (2006). Bioquímica. Ed. Médica Panamericana.