Prometafase: en la mitosis y en la meiosis
La prometafase es un estadio del proceso de división celular, intermedio entre la profase y la metafase. Se caracteriza por la interacción de los cromosomas en división con los microtúbulos que se encargarán de separarlos. Tanto en la mitosis como en la meiosis ocurre prometafase, pero con características distintas.
El objetivo claro de toda división celular es producir más células. Para conseguir esto, la célula originalmente debe duplicar su contenido de ADN; es decir, replicarlo. Además de ello, la célula debe separar estos cromosomas de manera que se cumpla el propósito particular de cada división de citoplasma.
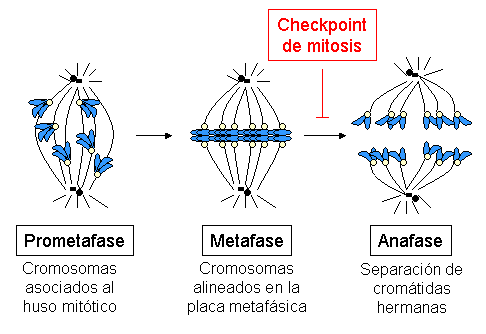
En la mitosis, el mismo número de cromosomas de la célula madre en las células hijas. En la meiosis I, la separación entre los cromosomas homólogos. En la meiosis II, la separación entre las cromátidas hermanas. Es decir, al final del proceso, obtener los cuatro productos meióticos esperados.
La célula maneja este complicado mecanismo a través del uso de componentes especializados como los microtúbulos. Estos son organizados por el centrosoma en la mayoría de los eucariotas. En otros, por el contrario, como las plantas superiores, funciona otro tipo de centro de organización de los microtúbulos.
Índice del artículo

Los microtúbulos son polímeros lineales de la proteína tubulina. Intervienen en casi todos los procesos celulares que involucran desplazamiento de alguna estructura interna. Son parte integral del citoesqueleto, cilios y flagelos.
En el caso de las células vegetales cumplen también un papel en la organización estructural interna. En estas células, los microtúbulos forman una especie de tapiz adosado a la cara interna de la membrana plasmática.
Esta estructura, que controla las divisiones celulares vegetales, se conoce con el nombre de organizacion cortical de los microtúbulos. En el momento de la división mitótica, por ejemplo, colapsan en un anillo central que será el futuro sitio de la placa central, en el plano donde se dividirá la célula.
Los microtúbulos están compuestos de alfa-tubulina y beta-tubulina. Estas dos subunidades forman un heterodímero, que es la unidad básica estructural de los filamentos de tubulina. La polimerización de los dímeros lleva a la formación de 13 protofilamentos en una organización lateral que da origen a un cilindro hueco.
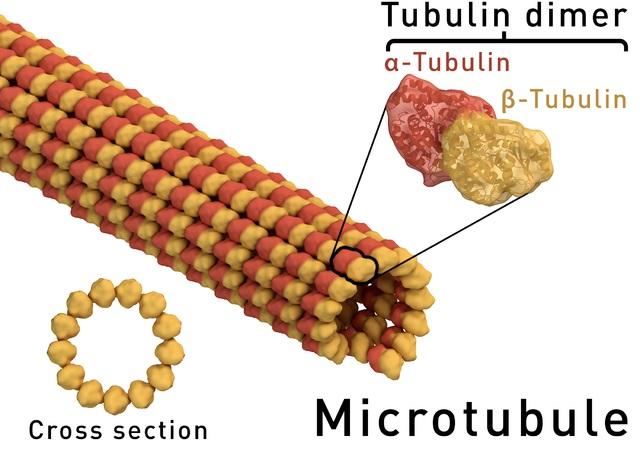
Los cilindros huecos de esta estructura son los microtúbulos, que por su propia composición muestran polaridad. Es decir, un extremo puede crecer por adición de heterodímeros, mientras que el otro puede sufrir sustracciones. En este último caso, el microtúbulo, en lugar de alargarse en esa dirección, se encoge.
Los microtúbulos se nuclean (es decir, comienzan a polimerizarse) y organizan en centros organizadores de los microtúbulos (COM). Los COM están asociados con los centrosomas durante las divisiones en células animales.
En las plantas superiores, que no presentan centrosomas, el COM está presente en sitios análogos, pero formado por otros componentes. En cilios y flagelos, el COM se ubica basalmente a la estructura motriz.
El desplazamiento de los cromosomas durante las divisiones celulares se consigue a través de los microtúbulos. Estos median la interacción física ente los centrómeros de los cromosomas y los COM.
Por reacciones de depolimerización orientadas, los cromosomas metafásicos eventualmente se moverán hacia los polos de las células en división.
La correcta segregación cromosomal mitótica es aquella que garantiza que cada célula hija reciba un complemento de cromosomas idénticos al de la célula madre.
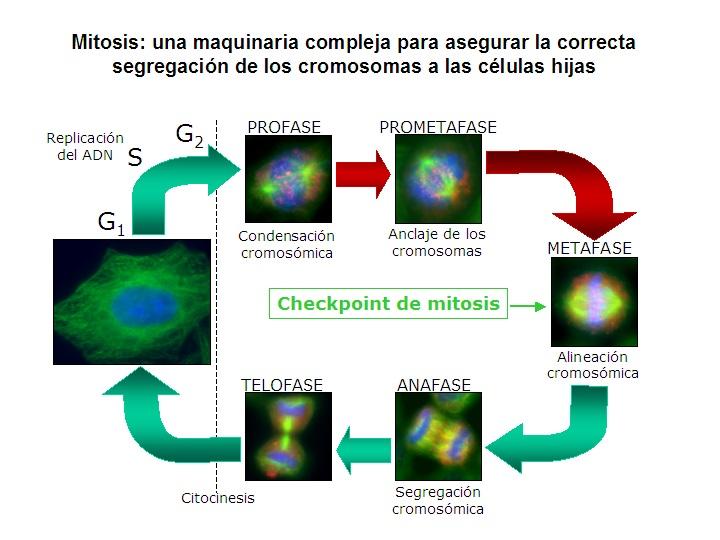
Esto significa que la célula debe separar a cada par de cromosomas duplicados en dos cromosomas individuales e independientes. Es decir, debe segregar las cromátidas hermanas de cada par homologo de todo el complemento de cromosomas de la célula madre.
En las mitosis abiertas, el proceso de desaparición de la envoltura nuclear es el rasgo distintivo de la prometafase. Esto permite que desaparezca el único obstáculo entre el MOC y los centrómeros de los cromosomas.
A partir de los MOC, se van polimerizando largos filamentos de microtúbulos que se van alargando hacia los cromosomas. Al encontrar un centrómero la polimerización cesa y se obtiene un cromosoma unido a un COM.
En la mitosis los cromosomas son dobles. Por lo tanto, también hay dos centrómeros, pero aun unidos en la misma estructura. Esto quiere decir que al finalizar el proceso de polimerización de los microtúbulos tendremos dos de ellos por cromosoma duplicado.
Un filamento unirá un centrómero a un COM, y otro a la cromátida hermana unida al COM opuesto al primero.
En las mitosis cerradas, el proceso es casi idéntico al anterior, pero con una gran diferencia; la envoltura nuclear no desaparece. Por lo tanto, el COM es interno y está asociado a la envoltura nuclear interna a través de la lámina nuclear.
En las mitosis semicerradas (o semiabiertas), la envoltura nuclear solo desaparece en los dos puntos opuestos donde por fuera del núcleo existe un COM mitótico.
Esto quiere decir que en estas mitosis los microtúbulos penetran hacia el interior del núcleo para poder movilizar a los cromosomas en pasos posteriores a la prometafase.
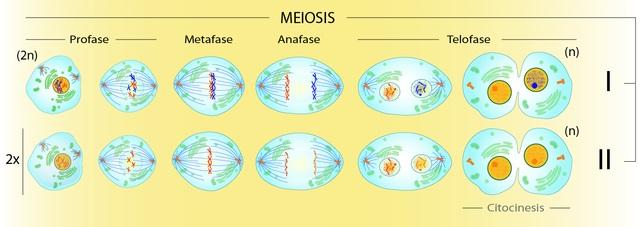
Como la meiosis implica la producción de cuatro células ‘n’ a partir de una célula ‘2n’, debe haber dos divisiones de citoplasma. Veámoslo así: al final de la metafase I habrá cuatro veces más cromátidas que centrómeros visibles al microscopio.
Tras la primera división, habrá dos células con del doble de cromátidas que de centrómeros. Solo al final de la segunda división citoplásmica se individualizarán todos los centrómeros y cromátidas. Habrá tantos centrómeros como haya cromosomas.
La proteína clave para que se realicen estas complejas interacciones intercromatínicas en la mitosis y la meiosis es la cohesina. Pero hay más complicaciones en la meiosis que en la mitosis. No es de extrañar, por lo tanto, que la cohesina meiótica sea distinta de la mitótica.
Las cohesinas permiten la cohesión de los cromosomas durante su proceso de condensación mitótico y meiótico. Además, permiten y regulan la interacción entre cromátidas hermanas en ambos procesos.
Pero en la meiosis también promueven algo que no ocurre en la mitosis: el apareamiento entre homólogos, y las consecuentes sinapsis. Estas proteínas son distintas en cada caso. Podríamos decir que la meiosis sin una cohesina que la distinga, no sería posible.
Mecanísticamente hablando, la interacción centrómero/COM es la misma en toda división celular. Sin embargo, en la prometafase I de la meiosis I la célula no va a separar cromátidas hermanas como lo hace en la mitosis.
Por el contrario, la tétrada meiótica posee cuatro cromátidas en una aparente juego doble de centrómeros. En esta estructura existe otra cosa no presente en la mitosis: quiasmas.
Los quiasmas, que son uniones físicas entre cromosomas homólogos, es lo que hace distinguir cuáles son los centrómeros que se deben segregar: los de los cromosomas homólogos.
Es así como en la prometafase I se forman conexiones entre centrómeros de los homólogos y COM en polos opuestos de la célula.
Esta prometafase II es más parecida a la prometafase mitótica que a la prometafase meiótica I. En este caso, los COM “lanzarán” microtúbulos a los centrómeros duplicados de las cromátidas hermanas.
Así, se producirán dos células con cromosomas individuales producto de una cromátida de cada par. Por lo tanto, se dará origen a células con el complemento cromosómico haploide la especie.
- Alberts, B., Johnson, A. D., Lewis, J., Morgan, D., Raff, M., Roberts, K., Walter, P. (2014) Molecular Biology of the Cell (6th Edition). W. W. Norton & Company, New York, NY, USA.
- Goodenough, U. W. (1984) Genetics. W. B. Saunders Co. Ltd, Philadelphia, PA, USA.
- Griffiths, A. J. F., Wessler, R., Carroll, S. B., Doebley, J. (2015). An Introduction to Genetic Analysis (11th ed.). New York: W. H. Freeman, New York, NY, USA.
- Ishiguro, K.-I. (2018) The cohesin complex in mammalian meiosis. Genes to Cells, doi: 10.1111/gtc.12652
- Manka, S. W., Moores, C. A. (2018) Microtubule structure by cryo-EM: snapshots of dynamic instability. Essays in Biochemistry, 62:737-751.