Sistema del complemento: vía clásica y alternativa, funciones, enfermedades
El sistema de complemento es un grupo integrado por más de treinta proteínas plasmáticas susceptibles al calor, que incrementan el efecto destructivo de microorganismos patógenos.
Recibe el nombre de “complemento” ya que se ha demostrado que complementa la acción de los anticuerpos en la destrucción de patógenos. Sin embargo, este es capaz también de llevar a cabo sus funciones en ausencia de anticuerpos. Por tanto, puede ser considerado como parte de los componentes del sistema inmune innato.
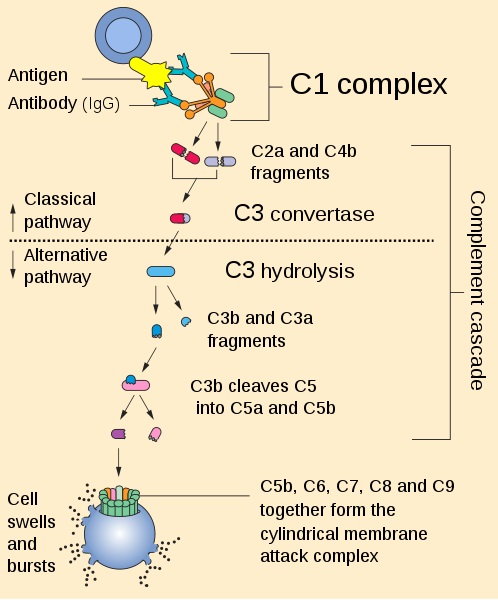
Su acción depende de la activación en serie (“cascada”) de las proteínas que lo integran, para así garantizar la ruptura de patógenos por medio de formación de poros en su membrana, el etiquetado (opsonización) para su destrucción por células fagocíticas y la neutralización de virus.
Índice del artículo
- 1 El sistema inmune: Inmunidad adaptativa e inmunidad innata
- 2 Sistema del complemento
- 3 ¿Cómo ocurre la activación del sistema de complemento?
- 4 El complemento puede ser activado por tres vías independientes
- 5 Funciones
- 6 Enfermedades relacionadas
- 7 Referencias
El sistema inmunitario es el sistema de defensa que tiene el organismo para defenderse del ataque de microorganismos capaces causar enfermedades.
Se encuentra compuesto por un conjunto de células, órganos y proteínas citoquinas que permanecen alerta ante la llegada de los patógenos. Una vez que los detectan, realizan el ataque contra estos a fin de garantizar su eliminación. Su metodología sería tal y como la harían los soldados de un cuartel, los cuales salen a la defensa siempre que se presenten situaciones de ataque o emergencia.
Como en todo sistema de defensa, el ataque que realizan requiere de tácticas, habilidades, destrezas y cooperación de sus componentes. Todo esto se engrana en una serie de pasos estratégicos que se conocen en conjunto como respuesta inmune.
La respuesta inmune se da en dos grandes fases separadas temporalmente: la respuesta inmune innata y la respuesta inmune adaptativa.
La respuesta inmune innata es la primera línea de defensa ante una infección provocada por la llegada de un organismo extraño.
Este tipo de respuesta inicial implica, por un lado, la acción de líneas de contención (la piel y las mucosas) que actúan como barreras impidiendo la entrada de patógenos. Por el otro, la acción de células que permanecen vigilantes en las capas más internas de la piel ante la entrada de los patógenos. Estos microorganismos pueden ‘colarse’ como consecuencia de una falla en las primeras barreras, como por ejemplo, un orificio o corte que exista en ellas.
Las células que actúan a este nivel son conocidas como fagocitos, los cuales se encargan de reconocer los microorganismos invasores, de fagocitarlos (devorarlos) y finalmente destruirlos en su citoplasma.
Aparte de esto, estas células se encargan de enviar señales a las células que participan en la segunda rama de respuesta a fin de eliminar eficientemente cualquier patógeno que logre superar la primera línea de respuesta.
Finalmente, los componentes celulares y no celulares que participan en este tipo de respuesta se encuentran presentes desde el nacimiento del organismo. Es decir, no dependen de la presencia de antígenos (patógenos o sustancias tóxicas extrañas).
Este tipo de respuesta, que acontece luego de que se han desencadenado los mecanismos efectores de la inmunidad innata, son llevadas a cabo por otras células conocidas como linfocitos.
Los linfocitos refuerzan los mecanismos de defensa de la inmunidad innata, al mismo tiempo que hacen que el sistema recuerde a los organismos invasores, por si acaso regresan.
Es decir, que ante una segunda invasión por un organismo extraño, estas últimas lo reconocen rápidamente facilitando su pronta eliminación. Estas respuestas suelen ser más rápidas que las primeras precisamente por su característica memoria inmunológica.
Finalmente, hay que mencionar que la inmunidad adaptativa se va desarrollando a lo largo de toda la vida de un organismo. Conforme este se va enfrentando a diferentes agentes infecciosos. Es decir, se adquiere.
Cuando estas células detectan un organismo por segunda vez desencadenan una línea de ataque celular y una humoral. La segunda involucra la liberación de anticuerpos, unas proteínas que neutralizan toxinas y marcan a los patógenos para su eliminación.
Los anticuerpos, a su vez, pueden activar a un grupo de proteínas que forman el sistema de complemento. Este último ayuda a destruir velozmente gérmenes y células ya infectadas.
El sistema de complemento es un conjunto de proteínas plasmáticas que se activan por la presencia de organismos patógenos.
Aunque esta activación depende en muchos casos de los anticuerpos (componentes de las respuestas adaptativas), también puede activarse en ausencia de ellos. Por tal razón, es considerado un componente importante de las respuestas innatas.
Las proteínas que integran este sistema son más de 30. Interactúan entre sí para complementar la acción de los anticuerpos y las células fagocíticas en la eliminación de patógenos.
Estas proteínas se han identificado con la letra “C” de complemento, y se forman por combinación de 9 proteínas (C1 a C9). Todas ellas son proteasas y se mantienen circulando vigilantes e inactivas por el cuerpo.
Una vez que se detecta la presencia de un microorganismo extraño, estas son activadas por la acción de otras proteasas, a fin de que salgan al ataque en defensa del organismo.
Ahora bien, dicha activación puede llevarse a cabo mediante tres vías diferentes: la vía clásica, la alternativa y la de las lectinas. Aunque, estas se diferencian en cómo se da la activación, todas coinciden en la formación de un complejo de ataque a la membrana del patógeno (MAC).
Este complejo se forma por la asociación de muchas proteínas en la cara externa de la membrana del patógeno que culmina con la formación de poros o huecos en esta.
La activación ocurre en los sitios donde ocurre la infección y es provocada por la presencia de los microorganismos invasores.
Durante la misma, todas las proteínas del complemento que se encuentran inicialmente inactivas son activadas en una reacción en cadena. Es decir, una vez que se ha activado una, esta última activa a la siguiente y así sucesivamente.
Las proteasas activas son generadas por el corte de la proteína precursora o zimógeno (forma inactiva). Esta última corta en dos a la siguiente activándola.
Entonces, la activación de un grupo pequeño de proteínas al principio de la cascada causa un incremento enorme en la activación de los zimógenos sucesivos (amplificación).
Dicha amplificación, ayuda a que el complejo de ataque a la membrana del patógeno se forme rápidamente. Esto promueve la apertura de poros que terminarán por romper a parásitos, bacterias y otros organismos capaces de provocar infección.
Aunque el objetivo final que se busca con la activación del complemento es siempre la formación del complejo de ataque a la membrana de patógenos, existen tres vías mediante las cuales puede lograrse este cometido. El inicio de cada una de ellas depende de la acción de moléculas diferentes.
Sin embargo, Todas convergen en la activación de la C3 convertasa, una proteína que fragmenta a la proteína C3 en C3a y C3b. Esta última se une a la membrana del patógeno y fractura a C5 en C5a y C5b. C5b se une también a la membrana y recluta a el resto de proteínas que se ensamblaran para dar origen al poro (C6, C7,C8 y C9).
Recibe este nombre por ser la primera vía en ser descrita. Constituye un punto de enlace entre los mecanismos de las respuestas innatas y las adaptativas ya que es activada por complejos de anticuerpos que se han unido previamente a la superficie del patógeno.
Esta comienza con la unión de C1q (la primera proteína de la cascada del complemento) a la membrana del microorganismo invasor. Esta unión se puede dar de tres formas diferentes:
– Directamente con componentes proteicos y no proteicos de la superficie de bacterias, como por ejemplo, el ácido lipoteicoico presente en bacterias grampositivas.
– A la proteína C reactiva, una proteína plasmática que se une a los residuos de fosfocolina presentes en polisacáridos de superficie bacteriana.
– A inmunocomplejos, formados por dos o más anticuerpos de los isotipos IgG o IgM que se han unido previamente al patógeno.
La activación por esta vía depende del reconocimiento de carbohidratos específicos expuestos en la superficie del patógeno por proteínas denominadas lectinas.
Las lectinas, son proteínas que solo interactúan con carbohidratos. Algunos ejemplos de estas son: la proteína MLB que se une específicamente a polisacáridos que contienen el azúcar manosa presente en la superficie de virus y bacterias, y las que reconocen solo residuos de N-acetilglucosamina presente en la pared bacteriana.
Esta vía se activa directamente por la unión de la proteína C3 (que genera C3b) ya activa a la superficie del patógeno.
Es importante saber que en ausencia de infecciones C3b se produce por esta vía en valores muy bajos. Estas limitadas cantidades de C3b se mantienen inactivas por la acción de una proteína conocida como factor H.
Solo cuando hay infección y C3 se une al patógeno, se evade el efecto regulador del factor H y esta se une a un segundo factor conocido como factor B. Este último se escinde por la acción del factor D y los productos se unen a la C3 presente ya en la membrana formando la C3 convertasa.
A partir de aquí se siguen los pasos de activación comunes a las tres vías.
Permite provocar la rápida destrucción de las células patógenas por medio de la formación de poros que destruyen velozmente la membrana de estas.
Mediante la unión de proteínas del complemento activadas marca a los patógenos para que sean reconocidos e ingeridos por células fagocíticas para su destrucción. Este proceso se conoce como opsonización.
Los fragmentos pequeños que se producen de la ruptura de los zimógenos actúan como quimioatrayentes que reclutan más fagocitos al lugar de infección.
Permite neutralizar virus invasores. Es decir, que los inactiva para que luego sean fagocitados y eliminados.
Deficiencias en la síntesis de proteínas del complemento así como factores que produzcan una activación no regulada de estas proteínas pueden dar lugar a numerosas enfermedades.
Las deficiencias son ocasionadas generalmente por errores genéticos que dan lugar a eventos de activación erróneos. Esto termina en insuficiencia en un incremento en la susceptibilidad a infecciones, enfermedades reumáticas y angioedema (edema en piel y mucosas).
La ausencia de regulación como por ejemplo, la ausencia del Factor H, pueden ocasionar un exceso de activación. Esto, termina en una inflamación descontrolada, producida por la lisis de células propias.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. 2002. Molecular Biology of the Cell, 4th edition. New York: Garland Science.
- McCulloch J, Martin SJ. Assays of cellular activity. 1994. Cellular Immunology, pp.95-113.
- Rich R, Fleisher T, Shearer W, Schroeder H, Frew A, Weyand C. 2012. Clinical Immunology, 4th edition. Canada: Elsevier.
- Sarma JV, Ward PA. The complement system. Cell and tissue research. 2011; 343(1), 227-235.
- Thomas J, Kindt Richard A. Goldsby Amherst College Barbara A. Osborne. Javier de León Fraga (Ed.). 2006. En Inmunología de Kuby sexta edición. pp. 37, 94-95.
- Trascasa L. Deficiencias del complemento. Diagnóstico de laboratorio. Presentación del registro español de deficiencias del complemento. Registro español de deficiencias del complemento. 2000; 19: 41-48.