Axonema: características y composición
El axonema es una estructura citoesqueletal interna de cilios y flagelos basada en microtúbulos y que le da movimiento a los mismos. Su estructura está constituida por una membrana plasmática que rodea un par de microtúbulos centrales y nueve pares de microtúbulos periféricos.
El axonema está ubicado afuera de la célula y se ancla al interior de esta por medio del cuerpo basal. Tiene 0,2 μm de diámetro y su longitud puede variar desde 5–10 μm en cilios hasta varios mm en el flagelo de algunas especies, aunque estos generalmente miden 50 –150 μm.
La estructura del axonema de cilios y flagelos es altamente conservativa en todos los organismos eucariotas, desde microalgas Chlamydomonas hasta el flagelo del espermatozoide humano.
Índice del artículo
- 1 Características
- 2 Estructura y composición
- 3 Excepciones al modelo “9+2” del axonema
- 4 Mecanismo de movimiento del axonema
- 5 Enfermedades relacionadas al axonema
- 6 Referencias
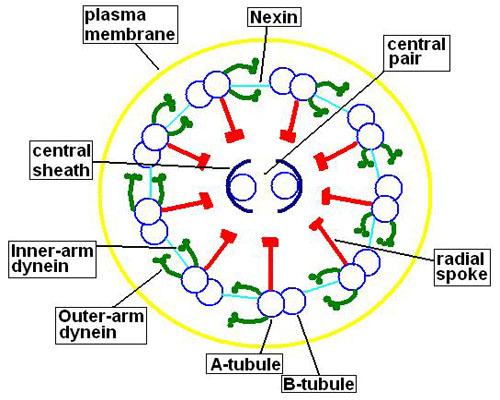
Los axonemas de la gran mayoría de los cilios y flagelos presentan una configuración conocida como “9+2”, es decir, nueve pares de microtúbulos periféricos rodeando un par central.
Los microtúbulos de cada par son diferentes en tamaño y composición, excepto el par central, que presenta ambos microtúbulos semejantes. Estos túbulos son estructuras estables capaces de resistir a rupturas.
Los microtúbulos presentan polaridad y todos tienen la misma disposición, con su extremo “+” ubicado hacia el ápice y el extremo “-“ ubicado basalmente.
Como ya señalamos, la estructura del axonema es de tipo 9+2. Los microtúbulos son estructuras cilíndricas largas, formados por protofilamentos. Los protofilamentos, a su vez, están constituidos por subunidades de proteínas denominadas tubulina alfa y tubulina beta.
Cada protofilamento posee una unidad de tubulina alfa en un extremo, mientras que el otro extremo posee una unidad de tubulina beta. El extremo con el terminal de tubulina beta recibe el nombre de extremo “+”, el otro extremo sería el extremo “-“. Todos los protofilamentos de un mismo microtúbulo están orientados con la misma polaridad.
Los microtúbulos contienen, además de las tubulinas, unas proteínas denominadas proteínas relacionadas con los microtúbulos (MAP, por sus siglas en inglés). De cada par de microtúbulos periféricos, el de menor tamaño (microtúbulo A) está compuesto de 13 protofilamentos.
El microtúbulo B posee solo 10 protofilamentos, pero es de mayor tamaño que el microtúbulo A. El par central de microtúbulos posee el mismo tamaño y cada uno de ellos está compuesto por 13 protofilamentos.
Este par central de microtúbulos está encerrado por la vaina central, de naturaleza proteica, la cual va a conectarse con los microtúbulos A periféricos por medio de los rayos radiales. Por su parte, los microtúbulos A y B de cada par se unen entre sí por una proteína denominada nexina.
De los microtúbulos A parte también un par de brazos formados por una proteína denominada dineina. Esta proteína es la responsable de utilizar la energía disponible en el ATP para lograr el movimiento de cilios y flagelos.
Externamente, el axonema está recubierto por una membrana ciliar o flagelar que posee la misma estructura y composición de la membrana plasmática de la célula.
Aunque la composición “9+2” del axonema está altamente conservada en la mayoría de las células eucariotas ciliadas y/o flageladas, existen algunas excepciones a este modelo.
En los espermatozoides de algunas especies, se pierde el par central de microtúbulos, dando lugar a una configuración “9+0”. El movimiento flagelar en estos espermatozoides no parece variar mucho del observado en axonemas con configuración normal, por lo cual se cree que estos microtúbulos no poseen una participación importante en el movimiento.
Este modelo de axonema ha sido observado en los espermatozoides de especies como el pez Lycondontis y de anélidos del género Myzostomum.
Otra configuración observada en los axonemas es la configuración “9+1”. En este caso se presenta un único microtúbulo central, en lugar de un par. En tales casos, el microtúbulo central está ampliamente modificado, presentando varias paredes concéntricas.
Este modelo de axonema ha sido observado en los gametos masculinos de algunas especies de gusanos planos. En estas especies, sin embargo, este modelo de axonema no se repite en otras células ciliadas o flageladas de los organismos.
Estudios del movimiento de los flagelos han demostrado que la flexión de estos ocurre sin que haya una contracción o acortamiento de los microtúbulos del axonema. Debido a ello, el citólogo Peter Satir ha propuesto un modelo de movimiento flagelar basado en el desplazamiento de los microtúbulos.
Según ese modelo, el movimiento se logra gracias al desplazamiento de un microtúbulo de cada par sobre su compañero. Este modelo es parecido al deslizamiento de las cadenas de miosina sobre la actina durante la contracción muscular. El movimiento ocurre en presencia de ATP.
Los brazos de dineina se encuentran anclados en el microtúbulo A de cada par, con los extremos dirigidos hacia el microtúbulo B. Al comienzo del movimiento, los brazos de dineina se adhieren al sitio de unión en el microtúbulo B. Luego, ocurre un cambio en la configuración de la dineina que impulsa al microtúbulo B hacia abajo.
La nexina mantiene ambos microtúbulos próximos entre sí. Posteriormente, los brazos de dineína se separan del microtúbulo B. Luego volverá a unirse para repetir el proceso. Este deslizamiento ocurre alternadamente entre un lado y otro del axonema.
Este desplazamiento en forma alternada en un lado y otro del axonema ocasiona que el cilio, o el flagelo, se doble primero hacia un lado y luego hacia el lado contrario. La ventaja del modelo de movimiento flagelar de Satir es que explicaría el movimiento del apéndice independientemente de la configuración del axonema de los microtúbulos del axonema.
Existen diversas mutaciones genéticas que pueden ocasionar un desarrollo anormal del axonema. Estas anormalidades pueden ser, entre otras, la carencia de uno de los brazos de dineina, ya sea el interno o el externo, de los microtúbulos centrales o de los rayos radiales.
En estos casos, se desarrolla un síndrome denominado síndrome de Kartagener, en el cual, las personas que lo padecen son infértiles debido a que los espermatozoides no son capaces de desplazarse.
Estos pacientes desarrollan además vísceras en posición invertida con relación a la posición normal; por ejemplo, el corazón ubicado en el lado derecho del cuerpo y el hígado en el izquierdo. Esta condición se conoce como situs inversus.
También son propensos los que padecen del síndrome de Kartagener de sufrir infecciones respiratorias y sinusuales.
Otra enfermedad relacionada con un desarrollo anormal del axonema es una enfermedad renal poliquística. En esta, se desarrollan múltiples quistes en los riñones que terminan por destruir al riñón. Tal enfermedad se debe a una mutación en los genes que codifican unas proteínas denominadas policistinas.
- M. Porter & W. Sale (2000). The 9 + 2 axoneme anchors multiple inner arm dyneins and a network of kinases and phosphatases that control motility. The Journal of Cell Biology.
- Axoneme. En Wikipedia. Recuperado de en.wikipedia.org.
- G. Karp (2008). Cell and molecular biology. Concepts and experiments. 5th Edition. John Wiley & Sons, Inc.
- S.L. Wolfe (1977). Biología de la célula. Ediciones Omega, S.A.
- T. Ishikawa (2017). Axoneme Structure from Motile Cilia. Cold Spring Harbor Perspectives in Biology.
- R.W. Linck, H. Chemes & D.F. Albertini (2016). The axoneme: the propulsive engine of spermatozoa and cilia and associated ciliopathies leading to infertility. Journal of Assisted Reproduction and Genetics.
- S. Resino (2013). El citoesqueleto: microtúbulos, cilios y flagelos. Recuperado de epidemiologiamolecular.com