Tilacoides: qué es, características, estructura, funciones
¿Qué son los tilacoides?
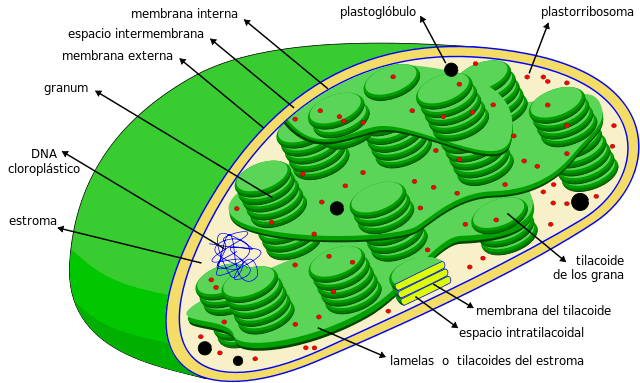
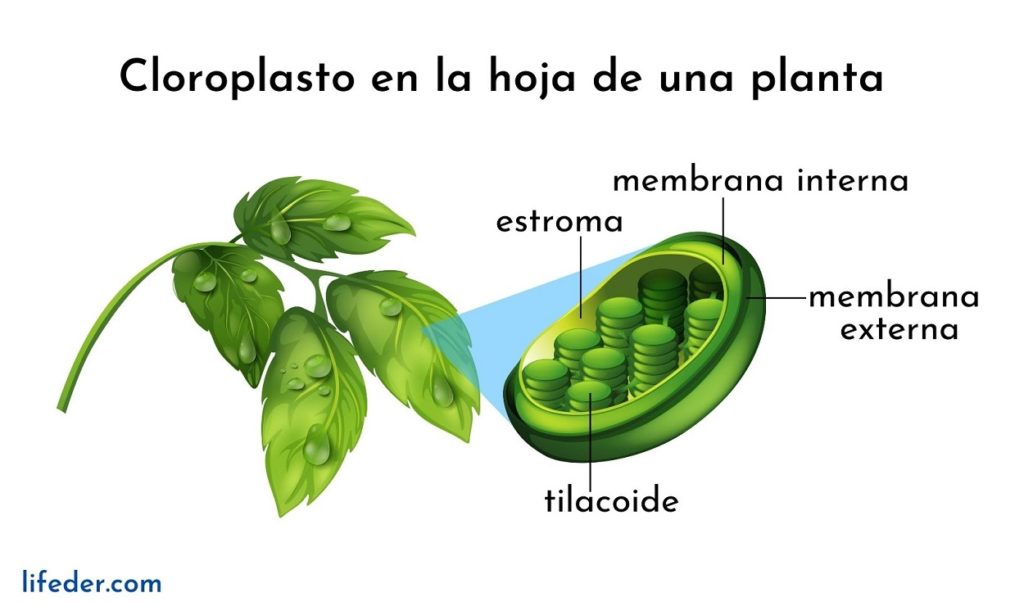
Los tilacoides son compartimientos en forma de sacos planos ubicados dentro de los cloroplastos en las células vegetales de las plantas, en las cianobacterias y algas. Suelen estar organizados en una estructura denominada grana —plural granum— y luce como una pila de monedas.
Los tilacoides son considerados el tercer sistema de membrana de los cloroplastos, aparte de la membrana interna y externa de dicha organela. La membrana de esta estructura separa al interior del tilacoide del estroma del cloroplasto, y posee una serie de pigmentos y proteínas involucrados en vías metabólicas.
En los tilacoides se dan reacciones bioquímicas indispensables para la fotosíntesis, proceso por el que las plantas toman luz solar y la transforman en carbohidratos. Específicamente, poseen la maquinaria necesaria anclada a su membrana para llevar a cabo la fase dependiente de luz solar, donde la luz es atrapada y convertida en energía (ATP) y NADPH.
Características de los tilacoides
– Los tilacoides son un sistema membranoso tridimensional interno de los cloroplastos. Los cloroplastos totalmente maduros poseen de 40 a 60 grana apiladas, con un diámetro comprendido entre 0,3 y 0,6 µm.
– El número de tilacoides que constituyen las granas varía ampliamente: desde menos de 10 sacos en plantas expuestas a suficiente luz solar, hasta más de 100 tilacoides en plantas que viven en ambientes con sombra extrema.
– Los tilacoides apilados se encuentran conectados entre sí formando un compartimiento continuo dentro del cloroplasto. El interior del tilacoide es un compartimiento bastante espacioso de naturaleza acuosa.
– La membrana de los tilacoides es indispensable para la fotosíntesis, ya que allí tiene lugar la primera etapa del proceso.
Estructura de los tilacoides
Los tilacoides son las estructuras que dominan dentro de un cloroplasto totalmente maduro. Si se visualiza un cloroplasto en el microscopio óptico tradicional, se podrán observar unas especies de granos.
Estos son los apilamientos de tilacoides; por ello, los primeros observadores de estas estructuras los denominaron “grana”.
Con ayuda del microscopio electrónico pudo ampliarse la imagen y se concluyó que la naturaleza de estos granos eran realmente tilacoides apilados.
La formación y estructura de la membrana del tilacoide depende de la formación del cloroplasto a partir de un plastidio aún no diferenciado, conocido como protoplastidio. La presencia de luz estimula la conversión a cloroplastos, y posteriormente la formación de los tilacoides apilados.
Membrana del tilacoide
En los cloroplastos y en las cianobacterias, la membrana del tilacoide no está en contacto con porción interna de la membrana plasmática. Sin embargo, la formación de la membrana del tilacoide empieza con la invaginación de la membrana interna.
En las cianobacterias y en ciertas especies de algas, los tilacoides están formados por una sola capa de lamelas. En contraste, existe un sistema más complejo encontrado en los cloroplastos maduros.
En este último grupo se pueden distinguir dos partes esenciales: el grana y la lamela del estroma. El primero consiste en pequeños discos apilados y la segunda se encarga de conectar estos apilamientos entre sí, formando una estructura continúa: el lumen del tilacoide.
Composición lipídica de la membrana
Los lípidos que conforman la membrana son altamente especializados y consisten en casi un 80 % de galactosil diacilglicerol: monogalactosil diacilglicerol y digalactosil diacilglicerol. Estos galactolípidos poseen cadenas altamente insaturadas, típicas de los tilacoides.
Del mismo modo, la membrana del tilacoide contiene en menor proporción lípidos como el fosfatidilglicerol. Los lípidos mencionados no se distribuyen de manera homogénea en ambas capas de la membrana; existe cierto grado de asimetría que pareciera contribuir en el funcionamiento de la estructura.
Composición proteína de la membrana
Los fotosistemas I y II son los componentes proteicos dominantes en esta membrana. Se encuentran asociados con el complejo del citocromo b6F y la ATP sintetasa.
Se ha encontrado que la mayoría de elementos del fotosistema II se ubica en las membranas de grana apilada, mientras que el fotosistema I se ubica mayormente en las membranas de tilacoides no apilados. Es decir, existe una separación física entre ambos fotosistemas.
Estos complejos incluyen proteínas integrales de membrana, proteínas periféricas, cofactores y una variedad de pigmentos.
Lumen del tilacoide
El interior del tilacoide consiste en una sustancia acuosa y espesa, cuya composición es distinta a la del estroma. Participa en la fotofosforilación, almacenando los protones que generarán la fuerza protón–motriz para la síntesis de ATP. En este proceso, el pH del lumen puede llegar a 4.
En el proteoma del lumen del organismo modelo Arabidopsis thaliana se han identificado más de 80 proteínas, pero sus funciones no han sido totalmente dilucidadas.
Las proteínas de lumen están involucradas en la regulación de la biogénesis de los tilacoides y en la actividad y recambio de las proteínas que forman los complejos fotosintéticos, especialmente el fotosistema II y la NAD(P)H deshidrogensa.
Funciones de los tilacoides
El proceso de fotosíntesis, vital para los vegetales, empieza en los tilacoides. La membrana que los delimita con el estroma de cloroplasto posee toda la maquinaria enzimática necesaria para que ocurran las reacciones fotosintéticas.
Etapas de la fotosíntesis
La fotosíntesis puede dividirse en dos grandes etapas: las reacciones lumínicas y las reacciones oscuras.
Como su nombre lo indica, las reacciones que pertenecen al primer grupo solo pueden proceder en presencia de luz, mientras que las del segundo grupo pueden surgir con o sin esta. Nótese que no es necesario que el ambiente esté “oscuro”, solo es independiente de la luz.
El primer grupo de reacciones, las “lumínicas”, ocurre en el tilacoide y puede resumirse de la siguiente manera: luz + clorofila + 12 H2O + 12 NADP+ + 18 ADP + 18 Pi à 6 O2 + 12 NADPH + 18 ATP.
El segundo grupo de reacciones ocurre en el estroma del cloroplasto y toma el ATP y el NADPH sintetizado en la primera etapa para reducir el carbono del dióxido de carbono a glucosa (C6H12O6). La segunda etapa puede resumirse en: 12 NADPH + 18 ATP + 6 CO2 à C6H12O6 + 12 NADP+ + 18 ADP + 18 Pi + 6 H2O.
Etapa dependiente de la luz
Las reacciones lumínicas involucran una serie de estructuras conocidas como fotosistemas, los cuales se encuentran en la membrana de los tilacoides y contienen en su interior unas 300 moléculas de pigmento, entre estas la clorofila.
Existen dos tipos de fotosistema: el primero posee un pico máximo de absorción de luz de 700 nanómetros y se conoce como P700, mientras que el segundo se denomina P680. Ambos están integrados en la membrana del tilacoide.
El proceso empieza cuando uno de los pigmentos absorbe un fotón y este “rebota” hacia otros pigmentos. Cuando una molécula de clorofila absorbe luz, un electrón salta y otra molécula lo absorbe. La molécula que perdió el electrón ahora está oxidada y posee carga negativa.
El P680 atrapa energía lumínica proveniente de la clorofila a. En este fotosistema, un electrón es arrojado a un sistema de mayor energía a un aceptor primario de electrones.
Este electrón cae al fotosistema I, pasando por la cadena transportadora de electrones. Este sistema de reacciones de oxidación y reducción se encarga de transferir protones y electrones de una molécula a otra.
En otras palabras, hay un flujo de electrones desde el agua al fotosistema II, al fotosistema I y al NADPH.
Fotofosforilación
Una porción de los protones generados por este sistema de reacciones se ubica en el interior del tilacoide (también llamado luz del tilacoide), creando un gradiente químico que genera una fuerza protón–motriz.
Los protones se mueven desde el espacio del tilacoide al estroma, siguiendo favorablemente el gradiente electroquímico; es decir, salen del tilacoide.
Sin embargo, el paso de los protones no es por cualquier sitio en la membrana, deben hacerlo a través de un sistema enzimático complejo llamado ATP sintetasa.
Este movimiento de protones hacia el estroma provoca la formación de ATP partiendo de ADP, proceso análogo al que ocurre en la mitocondria. La síntesis de ATP usando luz recibe el nombre de fotofosforilación.
Estas etapas mencionadas ocurren de manera simultánea: la clorofila del fotosistema II pierde un electrón y debe reemplazarlo con un electrón proveniente de la ruptura de una molécula de agua; el fotosistema I atrapa luz, se oxida y lanza un electrón que es atrapado por el NADP+.
El electrón perdido del fotosistema I es reemplazado por el resultante del fotosistema II. Estos compuestos serán usados en las reacciones posteriores de fijación del carbono, en el ciclo de Calvin.
Evolución
La evolución de la fotosíntesis como proceso liberador de oxígeno permitió la vida tal y como la conocemos.
Se plantea que la fotosíntesis se desarrolló hace unos miles de millones de años en el ancestro que dio origen a las cianobacterias actuales, a partir de un complejo fotosintético anóxico.
Se propone que la evolución de la fotosíntesis vino acompañada de dos sucesos indispensables: la creación del fotosistema P680 y la génesis de un sistema de membranas internas, sin conexión a la membrana celular.
Existe una proteína llamada Vipp1 esencial para la formación de los tilacoides. Efectivamente, esta proteína está presente en plantas, algas y cianobacterias, pero ausente en bacterias que realizan fotosíntesis anóxica.
Se cree que este gen pudo originarse por duplicación génica en el posible ancestro de las cianobacterias. Existe un solo caso de cianobacteria que es capaz de realizar fotosíntesis con oxígeno y no posee tilacoides: la especie Gloeobacter violaceus.