Herencia postmendeliana: teorías principales
La herencia postmendeliana comprende todas las ideas y/o trabajos que surgieron después de los estudios publicados por el naturalista religioso Gregorio Mendel en 1865, y que explican fenómenos o comportamientos genéticos diferentes a los descritos por este.
Gregorio Mendel es considerado el padre de la genética, puesto que estudiando cruces entre plantas de guisante, estableció las primeras leyes de la herencia hace más de 100 años. La primera ley, llamada “Ley de la segregación”, propone que los genes pueden existir en formas diferentes o alternativas.

Estas formas alternativas son llamadas “alelos” e influyen directamente en los caracteres fenotípicos de una especie. Los alelos pueden determinar el color del cabello, de los ojos, el tipo de cabello, etc.
En su primera ley, Mendel propuso que cada rasgo fenotípico transmitido de los padres a sus hijos está controlado por dos alelos, cada uno proveniente de un progenitor (uno de la “madre” y otro del “padre”).
Según esto, cuando los parentales aportan alelos diferentes, la progenie resulta con una mezcla, en donde uno de los alelos es dominante (el que se expresa en el fenotipo) y el otro es recesivo (no se ve, o está “latente”).
La segunda ley de Mendel se conoce como la “Ley de la herencia independiente”, ya que en sus cruces Mendel observó que el patrón de herencia que caracterizaba a los alelos que definían un rasgo no influía sobre el patrón de herencia de los alelos que definían a un rasgo diferente.
Índice del artículo
Desde que Gregor Mendel propuso sus famosas leyes hace más de 100 años se han dilucidado nuevos mecanismos para explicar la herencia de los caracteres, muchos de los cuales podría decirse que contradicen o difieren considerablemente de aquellos descritos por Mendel.
Ejemplo de los fenómenos descritos por la ciencia “moderna”, cuyos avances solo han sido posibles gracias al avance de las estrategias experimentales y los métodos de investigación, son la herencia ligada al sexo, la dominancia incompleta y la codominancia, entre otros.
La herencia ligada al sexo tiene que ver con los genes que se encuentra únicamente en los cromosomas sexuales. En los seres humanos, el sexo está determinado por los cromosomas X e Y. Las mujeres presentan dos copias de X (XX) y los hombres una de X y otra de Y (XY).

Los genes que se encuentran en cada uno de estos dos cromosomas son aquellos que contribuyen a aquellos rasgos que están ligados al sexo. Los genes en el cromosoma X están presentes en hombres y en mujeres, pero los genes de Y solo están en los hombres.
La herencia ligada al sexo fue observada y descrita por primera vez por William Bateson, uno de los primeros científicos en “redescubrir” los trabajos de Mendel y en aprovecharlos para realizar nuevos experimentos basados en sus conclusiones.
Bateson realizó cruces experimentales de gallinas con plumajes de diferentes colores, unas con plumas oscuras y lisas y otras con plumas lisas y con un patrón rayado.
Mediante estos cruces observó que, en la descendencia, los machos resultantes siempre tenían plumajes con un patrón rayado.
Solo en la tercera generación, cuando se cruzaba un macho rayado con una gallina de plumas oscuras y lisas, se obtenían algunos gallos con plumaje negro. Esta proporción resultante de los cruces desafiaba las proporciones propuestas por Mendel.
Herencia ligada al X
Después de los trabajos de Bateson, Thomas Henry Morgan encontró una explicación lógica para tan extraños resultados obtenidos por Bateson, atribuyendo la diferencia de coloración en las gallinas a que el patrón rayado era codificado por un alelo dominante que se encontraba en el cromosoma X.
En las gallinas, al contrario que en el ser humano, el sexo heterogamético (XY) corresponde a las hembras, entretanto el sexo homogamético corresponde a los machos (XX).
El alelo estudiado por Bateson y Morgan, entonces, era expresado por todos aquellos machos con plumaje rayado, pero cuando un macho heredaba dos cromosomas X con el alelo correspondiente a la coloración negra, entonces aparecían machos de plumaje oscuro y sin patrón rayado.
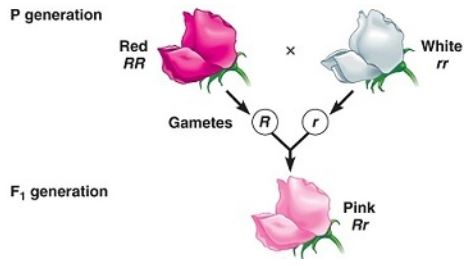
La dominancia incompleta es cuando el fenotipo que se observa en la descendencia que resulta del cruce de dos individuos homocigotos (uno dominante y otro recesivo) es un fenotipo “intermedio” entre el dominante y el recesivo.
Tiene que ver con la “incapacidad” del alelo “dominante” de enmascarar por completo la expresión del alelo recesivo, razón por la cual el fenotipo resultante corresponde a una mezcla de los rasgos codificados por ambos alelos, muchas veces descrito como un fenotipo “diluido” del dominante.
Por ejemplo, en las plantas Dondiego de la noche (Mirabilis jalapa) el color típico de los pétalos de las flores silvestres es rojo. Cuando una planta con pétalos rojos se cruza con una planta de línea pura con pétalos blancos se obtiene que toda la progenie es de pétalos rosas (rosados).
Cuando se cruzan dos individuos homocigotos dominantes y en la descendencia se observa un tercer fenotipo que corresponde a las dos características determinadas por los dos alelos parentales, hablamos de codominancia.
En este caso, siguiendo con un ejemplo similar al de las flores del ejemplo anterior, el “tercer fenotipo” no correspondería a flores rosadas, sino a una planta con flores blancas y con flores rojas en el mismo tallo, pues los dos alelos “codominan”, y ninguno enmascara al otro.
Muchas veces, sin embargo, la codominancia y la dominancia incompleta son difíciles de distinguir fenotípicamente, ya que ambas producen un fenotipo propio de un individuo que pareciera expresar dos alelos diferentes.
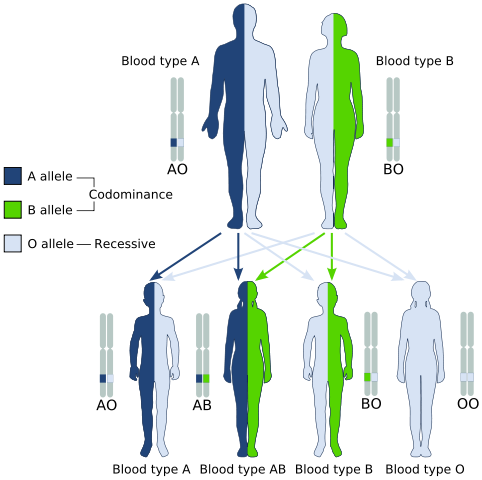
Sin embargo, genotípicamente los individuos que presentan alelos codominantes son diferentes a aquellos que presentan dominancia incompleta, ya que en un caso se expresan en la misma cantidad los dos alelos dominantes, y en el otro las proporciones son desiguales.
Un ejemplo clásico de codominancia es el tipo de grupo sanguíneo en los seres humanos, ya que los alelos que codifican para los antígenos A y B en las poblaciones humanas son codominantes.
Es decir, que cuando los alelos A y B se encuentran juntos en las células de un individuo, este tiene tipo de sangre AB.
Son alelos recesivos para algún carácter que, cuando se encuentran en forma homocigota, son letales para el individuo. Los individuos que presentan estos alelos no llegan a nacer, pues los mismos evitan el normal funcionamiento de la maquinaria molecular y evitan la traducción de los genes.
Muchos lugares del genoma de un individuo interactúan con otras partes del mismo genoma, es decir, que la modificación puntual de un alelo dentro del genoma puede condicionar de manera directa la expresión de otros alelos dentro de las células de un individuo.
Estas interacciones son difíciles de determinar y son clasificadas en dos tipos diferentes: los genes que interactúan en la misma ruta genética y aquellos que lo hacen en diferentes rutas genética.
Interacciones en diferentes rutas genéticas
Un ejemplo clásico y muy conocido de genes que interactúan en distintas rutas genéticas tiene que ver con los genes que determinan el color de la piel de las serpientes del maíz (Pantherophis guttatus), cuyo color común es un patrón repetitivo de mancha negras y anaranjadas.

Este fenotipo es producido por dos pigmentos diferentes que se encuentran bajo un estricto control genético. Un gen determina el color naranja y este tiene dos alelos: uno O+, que codifica para el pigmento y otro O-, que determina la ausencia total del pigmento naranja.
Otro gen diferente determina la presencia o la ausencia del pigmento negro, los alelos de este gen se conocen como B+, aquel que determina la presencia del pigmento negro y B-, que determina la ausencia del pigmento.
Ambos genes no están ligados y determinan independientemente el color de las serpientes, es decir, una serpiente con el genotipo O+/B+ es una serpiente normal con fenotipo naranja y negro, pero cualquier variación genotípica resultará en un fenotipo distinto.
Interacciones en la misma ruta genética
El ejemplo clásico de estos genes es el que se ha descrito para las flores “campanillas” (de la familia Campanulaceae), cuyo fenotipo “silvestre” corresponde a plantas con flores azules.
El color azul de las flores campanillas está determinado no por un gen, sino por una ruta metabólica donde varias enzimas, cada una codificada por un gen distinto, participan en la síntesis de un pigmento.
Así, existen fenotipos caracterizados por ser el resultado de la interacción de varios genes, cuyos productos participan en la misma ruta metabólica y donde la mutación o mala función de cualquiera de estos, rinde un fenotipo, en este caso, de flores blancas, despigmentadas.
- Ceccarelli, D. (2019). History of Inheritance.
- Griffiths, P., & Stotz, K. (2013). Genetics and philosophy: An introduction. Cambridge University Press.
- Lewin, B., Krebs, J. E., Goldstein, E. S., & Kilpatrick, S. T. (2014). Lewin’s Genes XI. Jones & Bartlett Publishers.
- McClearn, G. E., Vogler, G. P., & Plomin, R. (1996). Genetics and behavioral medicine. Behavioral Medicine, 22(3), 93-102.
- Richmond, M. L. (2008). William Bateson’s Pre-and Post-Mendelian Research Program in “Heredity and Development.”. A Cultural History of Heredity IV: Heredity in the Century of the Gene, 343, 213-242.