Herencia no mendeliana: patrones y ejemplos
Con “herencia no mendeliana” nos referimos a cualquier patrón de herencia en el cual los caracteres heredados no segreguen de acuerdo con lo estipulado por las leyes de Mendel.
En 1865, Gregor Mendel, considerado como el “padre de la genética”, realizó una serie de cruces experimentales con plantas de guisantes, cuyos resultados lo condujeron a proponer unos postulados (las leyes de Mendel) que buscaban dar una explicación lógica a la herencia de los caracteres entre padres e hijos.
Este astuto monje austríaco observó cuidadosamente la segregación de los genes parentales y su aparición en la descendencia como caracteres dominantes y recesivos. Además, determinó los patrones matemáticos que describían la herencia de una generación a otra y estos hallazgos fueron “ordenados” en forma de 3 leyes fundamentales:
– La ley de la dominancia
– La ley de la segregación de los caracteres y
– La ley de la distribución independiente.
Los aciertos y las deducciones de Mendel estuvieron ocultos durante muchos años, hasta su redescubrimiento a comienzos del siglo XX.
Por ese entonces, sin embargo, la comunidad científica conservó una posición un tanto escéptica respecto a estas leyes, ya que no parecían explicar los patrones de heredabilidad en cualquier especie animal o vegetal, especialmente en aquellos caracteres determinados por más de un locus.
Debido a esto, los primeros genetistas clasificaron los patrones hereditarios observados como “mendelianos” (aquellos que pudiesen ser explicados a través de la segregación de alelos simples, dominantes o recesivos, pertenecientes a un mismo locus) y “no mendelianos” (aquellos que no pudiesen ser explicados tan fácilmente).
Índice del artículo
La herencia mendeliana se refiere a un patrón hereditario que cumple con las leyes de la segregación y de la distribución independiente, según las cuales un gen heredado de cualquier parental segrega en los gametos con una frecuencia equivalente o, mejor dicho, con la misma probabilidad.
Los principales patrones de herencia mendeliana que han sido descritos para algunas enfermedades son: autosómico recesivo, autosómico dominante y ligado al cromosoma X, que se suman a los patrones de dominancia y recesividad descritos por Mendel.
Sin embargo, estos fueron postulados respecto a rasgos visibles y no a genes (debe tenerse en cuenta que algunos alelos pueden codificar rasgos que segregan como dominantes, mientras que otros pueden codificar los mismos rasgos, pero estos segregan como genes recesivos).
De lo anterior se deduce que la herencia no mendeliana consiste, sencillamente, en cualquier patrón hereditario que no cumpla con la norma en la cual un gen heredado de cualquier parental segregue en las células de la línea germinal con las mismas probabilidades y entre estos se incluyen:
– Herencia mitocondrial
– “Imprinting”
– Disomía uniparental
– Dominancia incompleta
– Codominancia
– Alelos múltiples
– Pleiotropía
– Alelos letales
– Rasgos poligénicos
– Herencia ligada al sexo
La ocurrencia de estas variaciones en los patrones hereditarios puede atribuírsele a las diversas interacciones que tienen los genes con otros componentes celulares, además del hecho de que cada uno está sujeto a regulación y variación en cualquiera de las etapas de transcripción, corte y empalme, traducción, plegamiento de proteínas, oligomerización, traslocación y compartimentación dentro de la célula y para su exportación.
En otras palabras, existen numerosas influencias epigenéticas que pueden modificar los patrones hereditarios de cualquier rasgo, lo que resulta en una “desviación” de las leyes de Mendel.
El ADN mitocondrial también transmite información de una generación a otra, tal y como lo hace el que está contenido en el núcleo de todas las células eucariotas. El genoma codificado en este ADN incluye los genes necesarios para la síntesis de 13 polipéptidos que forman parte de las subunidades de la cadena respiratoria mitocondrial, esenciales para los organismos con metabolismo aeróbico.
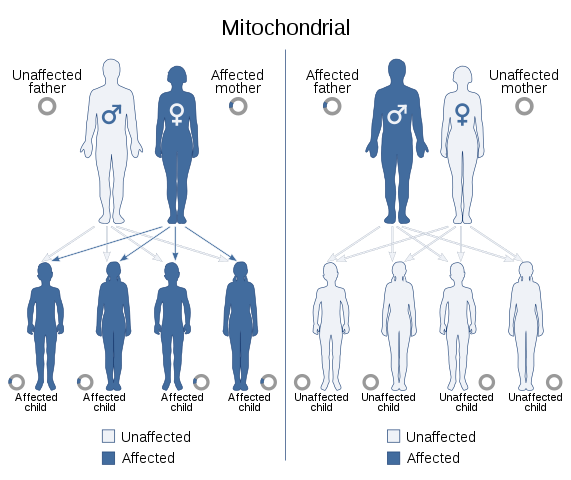
Aquellos rasgos que resultan de mutaciones en el genoma mitocondrial exhiben un patrón de segregación específica que ha sido llamado “herencia mitocondrial”, que usualmente ocurre a través de la línea materna, ya que el óvulo aporta el complemento total del ADN mitocondrial y ninguna mitocondria es aportada por el esperma.
La impronta genómica consiste en una serie de “marcas” epigenéticas que caracterizan a ciertos genes o regiones genómicas completas y que resultan del tránsito genómico del macho o de la hembra por el proceso de la gametogénesis.
Existen agrupamientos de impronta génica, los cuales consisten entre 3 y 12 genes distribuidos entre 20 y 3700 kilo bases de ADN. Cada agrupación posee una región conocida como región de control de la impronta, que exhibe modificaciones epigenéticas específicas provenientes de cada parental entre las que se incluyen:
– Metilación del ADN sobre alelos específicos en los residuos de citocina de pares CpG
– Modificación postraduccional de las histonas relacionadas con la cromatina (metilación, acetilación, fosforilación, etc., de las colas aminoacídicas de estas proteínas).
Ambos tipos de “marcas” modulan permanentemente la expresión de los genes sobre los que se encuentran, modificando sus patrones de transmisión a la siguiente generación.
A los patrones de herencia en los cuales la expresión de una enfermedad depende de los alelos específicos que son heredados de cualquiera de los dos padres se le conoce como el efecto de origen parental.
Este fenómeno es una excepción a la primera ley de Mendel, que establece que solo uno de los dos alelos presentes en cada parental es transmitido a la descendencia y, según las leyes cromosómicas de la herencia, solo uno de los cromosomas homólogos parentales puede ser transmitido a la siguiente generación.
Se trata de una excepción a la regla puesto que la disomía uniparental se trata de la herencia de ambas copias de un cromosoma homólogo de uno de los parentales. Este tipo de patrón hereditario no siempre muestra defectos fenotípicos, en vista de que mantiene las características numéricas y estructurales de los cromosomas diploides.
Este patrón hereditario consiste, fenotípicamente hablando, en una mezcla de los rasgos codificados por alelos que se combinan. En los casos de dominancia incompleta, aquellos individuos que son heterocigotos muestran una mezcla de los rasgos provenientes de los dos alelos que los controlan, lo que implica que la relación entre los fenotipos se modifica.
Describe los patrones hereditarios en los cuales los dos alelos que son transmitidos desde los padres hacia sus hijos se expresan simultáneamente en aquellos que poseen fenotipo heterocigoto, por lo que ambos se consideran “dominantes”.
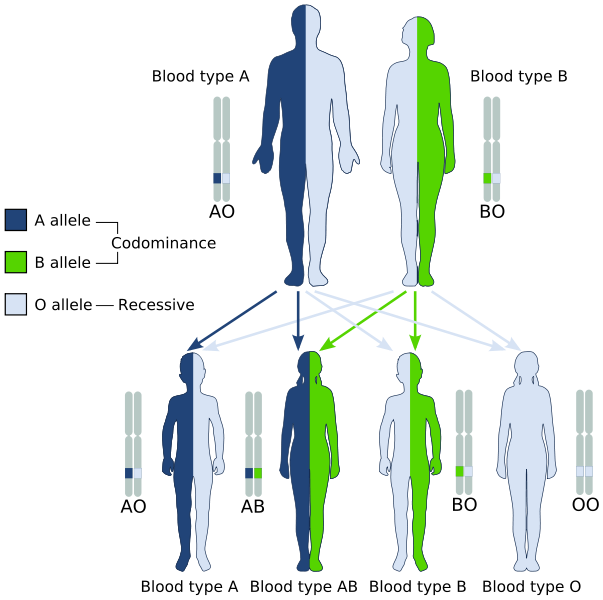
En otras palabras, el alelo recesivo no es “enmascarado” por la expresión del alelo dominante en la pareja alélica, sino que ambos se expresan y se observa, en el fenotipo, una mezcla de los dos rasgos.
Quizá una de las principales debilidades de la herencia mendeliana está representada por los rasgos que son codificados por más de un alelo, cosa que es bastante común en los humanos y en muchos otros seres vivos.
Este fenómeno hereditario incrementa la diversidad de los rasgos que son codificados por un gen y, adicionalmente, estos genes también pueden experimentar patrones de dominancia incompleta y codominancia además de la dominancia simple o completa.
Otra de las “piedras en el zapato” o de las “patas flojas” de las teorías hereditarias de Mendel tiene que ver con aquellos genes que controlan la aparición de más de un fenotipo o característica visible, como es el caso de los genes pleiotrópicos.
En sus trabajos, Mendel tampoco consideró la herencia de ciertos alelos que puede impedir la supervivencia de la descendencia cuando se encuentra en la forma homocigota o heterocigota; estos son los alelos letales.
Los alelos letales usualmente se relacionan con mutaciones o defectos en genes que son estrictamente necesarios para la supervivencia, que cuando son transmitidas a la siguiente generación (dichas mutaciones), dependiendo de la homocigosis o la heterocigosis de los individuos, resultan letales.
Existen características que están controladas por más de un gen (con sus alelos) y que, además, son fuertemente controladas por el ambiente. En los seres humanos esto es sumamente común y es el caso de rasgos como la estatura, el color de los ojos, del cabello y de la piel, así como el riesgo de padecimiento de algunas enfermedades.
En los seres humanos y muchos animales también existen rasgos que se encuentran en alguno de los dos cromosomas sexuales y que son transmitidos a través de la reproducción sexual. Muchos de estos rasgos se consideran “ligados al sexo” cuando son evidenciados en uno solo de los sexos, aunque los dos son físicamente capaces de heredar dichos rasgos.
La mayor parte de los rasgos ligados al sexo están asociados con algunas enfermedades y desórdenes recesivos.
Existe un desorden genético en los humanos conocido como el síndrome de Marfan, que es causado por una mutación en un solo gen que afecta simultáneamente el crecimiento y el desarrollo (estatura, visión y función cardíaca, entre otras).
Se trata de un caso considerado como un excelente ejemplo del patrón de herencia no mendeliana llamado pleiotropía, en el cual un solo gen controla diversas características.
Los desórdenes genéticos que resultan de mutaciones en el ADN mitocondrial presentan un sinnúmero de variaciones fenotípicas clínicas ya que ocurre lo que se conoce como heteroplasmia, donde diferentes tejidos tienen un porcentaje diferente de genoma mitocondrial mutante y, por lo tanto, presentan fenotipos diferentes.
Entre estos desórdenes están los síndromes de “agotamiento” de mitocondrias, que son un grupo de desórdenes autosómicos recesivos que se caracterizan por una reducción importante del contenido de ADN mitocondrial, lo que termina con sistemas de producción energética deficientes en aquellos órganos y tejidos más afectados.
Estos síndromes pueden deberse a mutaciones en el genoma nuclear que afectan a los genes nucleares involucrados en la síntesis de nucleótidos mitocondriales o en la replicación del ADN mitocondrial. Los efectos pueden ser evidenciados como miopatías, encefalopatías, defectos hepato-cerebrales o neuro-gastrointestinales.
- Gardner, J. E., Simmons, J. E., & Snustad, D. P. (1991). Principal of Genetic. 8 ‘“Edition. Jhon Wiley and Sons.
- Griffiths, A. J., Wessler, S. R., Lewontin, R. C., Gelbart, W. M., Suzuki, D. T., & Miller, J. H. (2005). An introduction to genetic analysis. Macmillan.
- Harel, T., Pehlivan, D., Caskey, C. T., & Lupski, J. R. (2015). Mendelian, Non-Mendelian, Multigenic Inheritance, and Epigenetics. In Rosenberg’s Molecular and Genetic Basis of Neurological and Psychiatric Disease (pp. 3-27). Academic Press.
- Silver, L. (2001). Non-Mendelian Inheritance.
- van Heyningen, V., & Yeyati, P. L. (2004). Mechanisms of non-Mendelian inheritance in genetic disease. Human molecular genetics, 13(suppl_2), R225-R233.