Síntesis de lípidos: tipos y sus principales mecanismos
La síntesis de lípidos consiste en una serie de reacciones enzimáticas por medio de las cuales se condensan hidrocarburos de cadenas cortas para formar moléculas de cadenas más largas que posteriormente pueden sufrir distintas modificaciones químicas.
Los lípidos son una clase de biomoléculas muy variadas sintetizadas por todas las células vivas y que están especializadas en múltiples funciones esenciales para el mantenimiento de la vida celular.
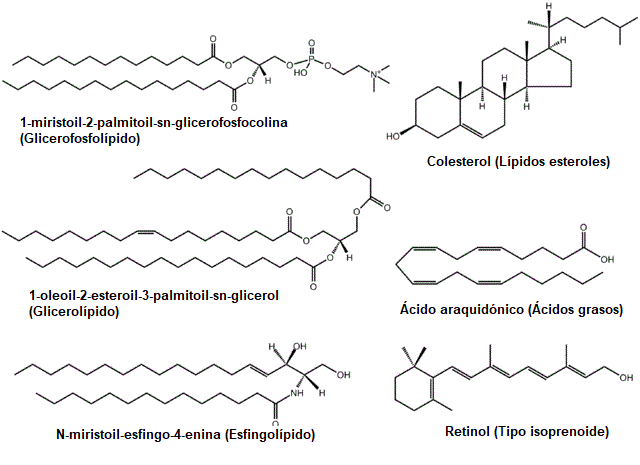
Los lípidos son los principales componentes de las membranas biológicas, hecho que los hace moléculas fundamentales para la existencia de las células como entidades aisladas de su entorno.
Algunos lípidos, además, tienen funciones especializadas como pigmentos, cofactores, transportadores, detergentes, hormonas, mensajeros intra- y extracelulares, anclas covalentes para proteínas de membrana, etc. Por tanto, la capacidad de sintetizar diferentes tipos de lípidos es crítica para la supervivencia de todos los organismos vivos.
Este gran grupo de compuestos se clasifica tradicionalmente en varias categorías o subgrupos: los ácidos grasos (saturados e insaturados), los glicéridos (fosfoglicéridos y glicéridos neutros), los lípidos no glicéridos (esfingolípidos (esfingomielinas y glucolípidos), esteroides y ceras) y los lípidos complejos (lipoproteínas).
Índice del artículo
Todas las secuencias de reacción de las rutas de biosíntesis de lípidos son endergónicas y reductivas. En otras palabras, todas utilizan ATP como fuente de energía y un transportador de electrones reducido, como el NADPH, como poder reductor.
A continuación, se describirán las principales reacciones de las rutas biosintéticas de los principales tipos de lípidos, es decir, de los ácidos grasos y eicosanoides, de los triacilgliceroles y fosfolípidos y de los esteroles (colesterol).
Los ácidos grasos son moléculas sumamente importantes desde el punto de vista lipídico, pues forman parte de los lípidos más relevantes en las células. Su síntesis, al contrario de lo que muchos científicos pensaban durante los primeros estudios al respecto, no consiste en la ruta inversa de su β-oxidación.
De hecho, esta ruta metabólica ocurre en compartimentos celulares diferentes y requiere de la participación de un intermediario de tres átomos de carbono conocido como malonil-CoA, que no es necesario en la oxidación.
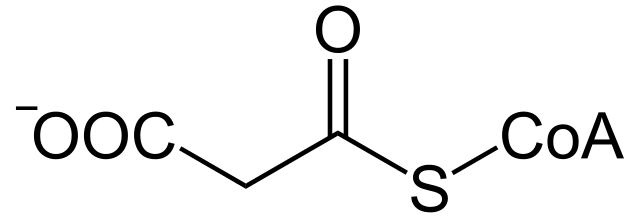
Además, se relaciona estrechamente con los grupos sulfhidrilo de unas proteínas conocidas como transportadoras de grupos acilo (ACP, del inglés Acyl Carrier Proteins).
En líneas generales, la síntesis de ácidos grasos, especialmente la de los de cadena larga, es un proceso secuencial donde se repiten cuatro pasos en cada “vuelta”, y durante cada vuelta se produce un grupo acilo saturado que es el sustrato para la siguiente, que implica otra condensación con una nueva molécula de malonil-CoA.
En cada vuelta o ciclo de reacción la cadena del ácido graso se extiende dos carbonos, hasta que alcanza una longitud de 16 átomos (palmitato), luego de lo cual abandona el ciclo.
Formación de malonil-CoA
Este intermediario de tres átomos de carbono es formado irreversiblemente a partir de acetil-CoA gracias a la acción de una enzima acetil-CoA carboxilasa, la cual posee un grupo prostético de biotina que está unido covalentemente a la enzima y que participa en esta catálisis en dos pasos.
En esta reacción, un grupo carboxilo derivado de una molécula de bicarbonato (HCO3-) es transferida a la biotina de forma ATP-dependiente, donde el grupo biotinil cumple la función de “transportador temporal” de la molécula mientras lo transfiere al acetil-Coa, produciendo malonil-CoA.
En la secuencia de síntesis de los ácidos grasos el agente reductor utilizado es el NADPH y los grupos activadores son dos grupos tiol (-SH) que forman parte de un complejo multienzimático llamado ácido graso sintasa, que es la que tiene mayor importancia en la catálisis sintética.
En los vertebrados, el complejo ácido graso sintasa es parte de una sola cadena polipeptídica de gran tamaño, en la cual están representadas las 7 actividades enzimáticas características de la ruta de síntesis, así como la actividad hidrolítica necesaria para liberar los intermediarios al final de la síntesis.

Las 7 actividades enzimáticas de este complejo son: proteína transportadora de grupo acilo (ACP), acetil-CoA-ACP transacetilasa (AT), β-cetoacil-ACP sintasa (KS), malonil-CoA-ACP transferasa (MT), β-cetoacil-ACP reductasa (KR), β-hidroxiacil-ACP deshidratasa (HD) y enoil-ACP reductasa (ER).
Antes de que puedan producirse las reacciones de condensación para ensamblar la cadena de ácido graso, los dos grupos tiol en el complejo enzimático se “cargan” con los grupos acilo: en primer lugar, un acetil-CoA es transferido al grupo -SH de una cisteína en la parte β-cetoacil-ACP sintasa del complejo, reacción catalizada por la enzima acetil-CoA-ACP transacetilasa (AT).
Posteriormente, un grupo malonil es transferido desde una molécula de malonil-CoA hacia el grupo -SH de la parte transportadora de grupo acilo (ACP) del complejo enzimático, reacción catalizada por una enzima malonil-CoA-ACP transferasa (MT), que también forma parte del complejo ácido graso sintasa.
La secuencia de cuatro reacciones por cada “vuelta” del ciclo de reacción es como sigue:
- Condensación: los grupos acetil y malonil “cargados” en la enzima se condensan para formar una molécula de acetoacetil-ACP, que está unida a la porción ACP a través de un grupo -SH. En este paso se produce una molécula de CO2 y es catalizado por la β-cetoacil-ACP sintasa (el grupo acetilo ocupa la posición de “metilo terminal” del complejo acetoacetil-ACP).
- Reducción del grupo carbonilo: el grupo carbonilo en la posición C3 del acetoacetil-ACP es reducido para formar D-β-hidroxibutiril-ACP, reacción catalizada por la β-cetoacil-ACP reductasa, que emplea NADPH como donador de electrones.
- Deshidratación: los carbonos C2 y C3 del D-β-hidroxibutiril-ACP son desprovistos de moléculas de agua, formándose un doble enlace que termina con la producción del nuevo compuesto trans-∆2-butenoil-ACP. Este proceso es mediado por una enzima β-hidroxiacil-ACP deshidratasa (HD).
- Reducción del doble enlace: el doble enlace del compuesto formado en el paso de deshidratación es saturado (reducido) para dar lugar a butiril-ACP mediante la reacción catalizada por la enzima enoil-ACP reductasa (ER), que también emplea NADPH como agente reductor.
Las reacciones de síntesis ocurren hasta cuando se forma una molécula de palmitato (16 átomos de carbono), el cual es hidrolizado del complejo enzimático y liberado como posible precursor para ácidos grasos de cadenas de mayor longitud, los cuales son producidos por unos sistemas de elongación de ácidos grasos ubicados en la porción lisa del retículo endoplásmico y en la mitocondria.
Las demás modificaciones que pueden sufrir estas moléculas como las desaturaciones, por ejemplo, son catalizadas por enzimas diferentes, que generalmente ocurren en el retículo endoplásmico liso.
Los eicosanoides son lípidos celulares que tienen funciones como moléculas mensajeras de “corto alcance”, producidas por algunos tejidos para comunicarse con las células de sus tejidos vecinos. Estas moléculas son sintetizadas a partir de ácidos grasos poliinsaturados de 20 átomos de carbono.
Prostaglandinas
En respuesta a un estímulo hormonal, la enzima fosfolipasa A ataca fosfolípidos de membrana y libera araquidonato desde el carbono 2 del glicerol. Este compuesto es convertido en prostaglandinas gracias a una enzima del retículo endoplásmico liso con actividad bifuncional: la ciclooxigenasa (COX) o prostaglandina H2 sintasa.
Tromboxanos
Las prostaglandinas pueden ser convertidas en tromboxanos gracias a la tromboxano sintasa presente en las plaquetas (trombocitos) sanguíneos. Estas moléculas participan en los pasos iniciales de la coagulación sanguínea.
Los ácidos grasos son moléculas fundamentales para la síntesis de otros compuestos más complejos en las células, como es el caso de los triacilgliceroles o de los lípidos glicerofosfolípidos de membrana (procesos que dependen de las necesidades metabólicas celulares).
Los animales producen triacilgliceroles y glicerofosfolípidos a partir de dos precursores comunes: el acil graso-CoA y el L-glicerol 3-fosfato. Los acil graso-CoA son producidos por las enzimas acil-CoA sintetasas que participan en la β-oxidación, mientras que el L-glicerol 3-fosfato es obtenido a partir de la glucólisis y por acción de dos enzimas alternativas: glicerol 3-fosfato deshidrogenasa y glicerol quinasa.
Los triacilgliceroles son formados por la reacción entre dos moléculas de acil graso-CoA y una molécula de diacilglicerol 3-fosfato; estas reacciones de transferencia son catalizadas por acil transferasas específicas.
En esta reacción se produce, inicialmente, ácido fosfatídico, el cual es desfosforilado por una enzima ácido fosfatídico fosfatasa para producir 1,2-diacilglicerol, que es capaz, nuevamente, de aceptar una tercera molécula de acil graso-CoA, produciéndose triacilglicerol.
Los fosfolípidos son moléculas sumamente variables, pues muchos diferentes pueden formarse por la combinación de ácidos grasos y grupos “cabeza” diferentes con el esqueleto de glicerol (glicerofosfolípidos) o de esfingosina (esfingolípidos) que los caracterizan.
El ensamblaje general de estas moléculas requiere de la síntesis del esqueleto de glicerol o esfingosina, de la unión con los ácidos grasos correspondientes, bien sea por esterificación o amidación, la adición de un grupo “cabeza” hidrofílico a través de un enlace fosfodiéster y, si es necesaria, la alteración o intercambio de estos últimos grupos.
En los eucariotas este proceso ocurre en el retículo endoplásmico liso y también en la membrana mitocondrial interna, donde pueden permanecer indefinidamente o desde donde pueden ser traslocados a otros lugares.
Pasos de reacción
Los primeros pasos de la reacción de síntesis de los glicerofosfolípidos son equivalentes a los de la producción de triacilgliceroles, pues una molécula de glicerol 3-fosfato es esterificada a dos moléculas de ácido graso en los carbonos 1 y 2, formando ácido fosfatídico. Es común hallar fosfolípidos que tienen ácidos grasos saturados en el C1 e insaturados en el C2 del glicerol.
El ácido fosfatídico también puede ser producido por fosforilación de una molécula de diacilglicerol ya sintetizada o “reciclada”.
Los grupos polares “cabeza” de estas moléculas se forman a través de enlaces fosfodiéster. Lo primero que debe ocurrir para que este proceso se dé correctamente es la “activación” de uno de los grupos hidroxilo que participa en el proceso mediante su unión a un nucleótido como la citidina difosfato (CDP), que es desplazada nucleofílicamente por el otro grupo hidroxilo que participa en la reacción.
Si esta molécula se une al diacilglicerol, entonces se forma CDP-diacilglicerol (la forma “activada” del ácido fosfatídico), pero esto puede ocurrir también sobre el grupo hidroxilo del grupo “cabeza”.
En el caso de la fosfatidilserina, por ejemplo, el diacilglicerol es activado por condensación de la molécula de ácido fosfatídico con una molécula de citidina trifosfato (CTP), formándose CDP-diacilglicerol y eliminándose un pirofosfato.
Si se desplaza una molécula de CMP (citidina monofosfato) por un ataque nucleofílico del hidroxilo de la serina o del hidroxilo en el carbono 1 del glicerol 3-fosfato se producen fosfatidilserina o fosfatidilglicerol 3-fosfato, del cual puede liberarse el monoéster de fosfato y producir fosfatidilglicerol.
Ambas moléculas producidas de esta forma sirven como precursores para otros lípidos de membrana, que muchas veces comparten las rutas biosintéticas entre sí.
El colesterol es una molécula esencial para los animales que puede ser sintetizada por sus células, por lo que no es fundamental en la dieta diaria. Esta molécula de 27 átomos de carbono es producida a partir de un precursor: el acetato.
Esta compleja molécula se forma a partir de acetil-CoA en cuatro etapas principales:
- Condensación de tres unidades de acetato para formar el mevalonato, una molécula intermediara de 6 carbonos (primero se forma una molécula de acetoacetil-CoA con dos acetil-CoA (enzima tiolasa) y después otra de β-hidroxi-β-metilglutaril-CoA (HMG-CoA) (enzima HMG-CoA sintetasa). El mevalonato se forma a partir de la HMG-CoA y gracias a la enzima HMG-CoA reductasa.
- Conversión de mevalonato en unidades de isopreno. Primero se transfieren 3 grupos fosfato desde 3 moléculas de ATP al mevalonato. Uno de los fosfatos se pierde junto con el grupo carbonilo adyacente y se forma ∆3-isopentenil pirofosfato, el cual es isomerizado para producir dimetilalil pirofosfato
- Polimerización o condensación de 6 unidades de isopreno de 5 átomos de carbono para formar el escualeno de 30 átomos de carbono (una molécula lineal).
- Ciclación del escualeno para formar los 4 anillos del núcleo esteroideo del colesterol y los cambios químicos subsecuentes: oxidaciones, migración y eliminación de grupos metilo, etc., lo que rinde colesterol.
- Garrett, R. H., & Grisham, C. M. (2001). Principles of biochemistry: with a human focus. Brooks/Cole Publishing Company.
- Murray, R. K., Granner, D. K., Mayes, P. A., & Rodwell, V. W. (2014). Harper’s illustrated biochemistry. Mcgraw-Hill.
- Nelson, D. L., Lehninger, A. L., & Cox, M. M. (2008). Lehninger principles of biochemistry. Macmillan.
- Jacquemyn, J., Cascalho, A., & Goodchild, R. E. (2017). The ins and outs of endoplasmic reticulum‐controlled lipid biosynthesis. EMBO reports, 18(11), 1905-1921.
- Ohlrogge, J., & Browse, J. (1995). Lipid biosynthesis. The Plant Cell, 7(7), 957.