Inervación del corazón: organización y actividad tónica
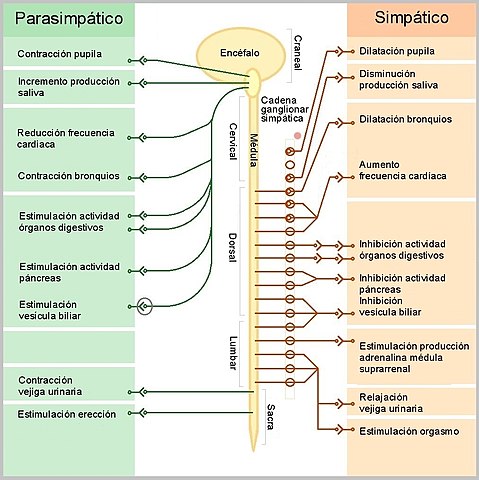
La inervación del corazón se organiza en la inervación simpática y en la parasimpática. Como cualquier otro órgano, el corazón recibe una inervación cuyas fibras se catalogan como pertenecientes al sistema nervioso autónomo (SNA), una de las dos divisiones del sistema nervioso periférico y encargada de mediar la sensibilidad y controlar la actividad visceral del organismo.
A pesar de tratarse de un órgano muscular estriado, muy semejante al músculo esquelético, el corazón no recibe inervación de la otra división del sistema periférico que media la sensibilidad somática y la actividad de los músculos que produce los desplazamientos de las articulaciones.
Cualquier proceso contráctil en un músculo esquelético necesita de una excitación inducida por una fibra nerviosa motora somática. El corazón, por su parte, no necesita ser excitado por nada externo a él mismo, puesto que tiene la capacidad de generar espontáneamente sus propias excitaciones.
Así pues, una de las características destacadas de la inervación autonómica cardíaca está representada por el hecho de que ella no es determinante de la actividad contráctil del corazón, que puede continuar después de su denervación, sino que ejerce una función moduladora de la misma.
Organización
La porción eferente o motora del sistema nervioso autónomo está organizada en dos componentes: el simpático y el parasimpático, sistemas constituidos por vías que conectan neuronas en el sistema nervioso central con células efectoras viscerales del organismo sobre las cuales ejercen efectos antagónicos.
Cada una de esas vías es una cadena de dos neuronas:
- Una preganglionar, cuyo cuerpo está en el sistema nervioso central y cuyo axón termina en un ganglio autonómico periférico, en el cual hace sinapsis con el cuerpo neuronal de una segunda neurona.
- La posganglionar, cuyo axón termina sobre el efector visceral.
Inervación simpática
Las células preganglionares simpáticas destinadas al corazón se originan en conglomerados celulares ubicados en las astas laterales de la médula espinal, en los segmentos torácicos T1-T5. Conglomerados celulares que constituyen en conjunto un “centro simpático cardioacelerador espinal”.
Sus axones representan fibras preganglionares que se dirigen a la cadena ganglionar simpática; especialmente a los ganglios cervicales superior, medio e inferior, en donde conectan con las neuronas posganglionares, cuyos axones se distribuyen con los nervios cardíacos superior, medio e inferior.
De estos tres nervios, el medio parece ser el que mayor influencia ejerce sobre las funciones cardíacas, porque el superior está destinado a las grandes arterias de la base del corazón y el inferior parece conducir información sensorial o aferente.
Un detalle más de la organización de la inervación simpática cardíaca es el que las fibras simpáticas derechas parecen terminar principalmente sobre el nodo sinoauricular, mientras que las izquierdas influyen sobre el nodo aurículo-ventricular, el sistema de conducción y el miocardio contráctil.
Acciones del simpático sobre el corazón
El sistema nervioso simpático ejerce una acción positiva sobre todas las funciones cardíacas, incrementando la frecuencia cardíaca (cronotropismo +), la fuerza de contracción (inotropismo +), la conducción de la excitación (dromotropismo +) y la velocidad de relajación (lusotropismo +).
Todas esas acciones se ejercen mediante la liberación de noradrenalina (NA) a nivel de las terminaciones posganglionares simpáticas sobre las células de los nodos cardíacos, del sistema de conducción o sobre los miocitos contráctiles auriculares y ventriculares.
Las acciones de la noradrenalina se desencadenan cuando este neurotransmisor se une a unos receptores adrenérgicos tipo β1 ubicados en las membranas de las células cardíacas y acoplados a una proteína Gs. Esta es una proteína con tres subunidades (αsβγ) que cuando está inactiva tiene GDP unido a su subunidad αs.
La interacción noradrenalina-receptor β1 hace que la subunidad αs libere su GDP y lo cambie por GTP; al hacerlo, se separa del componente βγ y activa a la enzima membranal adenilciclasa, la cual produce adenosín monofosfato cíclico (AMPc) como segundo mensajero que activa a la proteín quinasa A (PKA).
La actividad fosforilante de la PKA es en último término la responsable de todas las acciones estimuladoras que las fibras simpáticas ejercen sobre el corazón, e incluye la fosforilación de canales de Ca++, de la troponina I y del fosfolamban.
La acción sobre los canales de Ca++ favorece los incrementos de frecuencia cardíaca, de fuerza contráctil y de velocidad de conducción. Los efectos sobre la troponina I y sobre el fosfolamban aceleran el proceso de relajación del músculo cardíaco.
La fosforilación de la troponina I hace que esta proteína acelere el proceso de liberación del Ca++ de la troponina C para que la relajación ocurra más rápido. El fosfolamban inhibe naturalmente a la bomba que reintroduce el Ca++ al retículo sarcoplásmico para terminar la contracción, inhibición que se reduce cuando aquel se fosforila.
Inervación parasimpática
La inervación parasimpática del corazón cursa con el nervio vago y sus componentes tienen una organización de cadenas bineuronales similares a las del simpático, con neuronas preganglionares cuyos cuerpos se encuentran en el núcleo motor dorsal del vago en el bulbo, en el piso del cuarto ventrículo.
Debido a los efectos reductores de la actividad cardíaca que dichas neuronas ejercen sobre el corazón, han recibido en conjunto el nombre de “centro cardioinhibidor bulbar”. Sus fibras se separan del tronco vagal en el cuello y se entremezclan luego con las fibras simpáticas cardíacas formando un plexo.
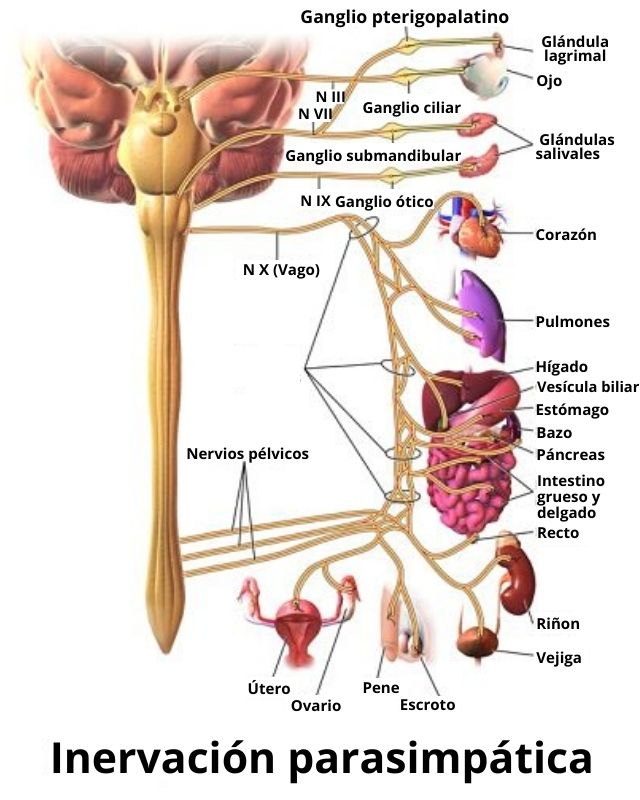
Los ganglios parasimpáticos se encuentran en las cercanías del corazón y las fibras posganglionares del lado derecho terminan fundamentalmente en el nodo sinoauricular, marcapaso natural del corazón, y las izquierdas en el nodo auriculoventricular y en los miocitos contráctiles auriculares.
Acciones del parasimpático sobre el corazón
La actividad parasimpática dirigida al corazón se traduce en un efecto negativo sobre algunas funciones cardíacas como disminución de la frecuencia (inotropismo -), de la velocidad de conducción en el nodo AV (dromotropismo -) y reducción de la fuerza contráctil de las aurículas (inotropismo auricular -).
La escasa o incluso inexistente inervación del parasimpático sobre el miocardio ventricular descarta un efecto inotrópico negativo de esta división autonómica sobre la fuerza contráctil de este músculo.
Las mencionadas acciones vagales sobre el corazón se ejercen mediante la liberación de acetilcolina (ACh) a nivel de las terminaciones posganglionares parasimpáticas sobre las células de los nodos cardíacos y los miocitos contráctiles auriculares.
Las acciones de la acetilcolina se desencadenan cuando ella se une a receptores colinérgicos muscarínicos tipo M2 ubicados en las membranas de las células mencionadas y acoplados a una proteína Gi. Esta posee tres subunidades (αiβγ) y cuando está inactiva tiene GDP unido a su subunidad αi.
La interacción acetilcolina-receptor M2 libera la subunidad αi. Esta inhibe a la adenilciclasa, se produce menos AMPc y se reduce la actividad de la PKA y la fosforilación de los canales de Ca++, efectos contrarios a los de la NA liberada por el simpático. El componente βγ activa una corriente de K+ (IKACh).
La reducción de la fosforilación de los canales de Ca++ reduce la corriente despolarizante de este ión, mientras que la aparición de la corriente IKACh introduce una corriente hiperpolarizante que se opone a la despolarización espontánea que produce los potenciales de acción (PA) en las células nodulares.
La reducción de la corriente de Ca++ despolarizante combinada con el incremento de la corriente de K+ hiperpolarizante enlentece el proceso de despolarización espontánea que va llevando automáticamente el potencial de membrana hasta el nivel umbral al cual se dispara el potencial de acción.
Este efecto puede llegar a ser de tal magnitud que una estimulación intensa del nervio vago puede llegar a detener el corazón, por desaparición de los potenciales de acción de las células marcapaso o por un bloqueo total del nodo aurículo-ventricular que no deja pasar los potenciales de acción de la aurícula derecha a los ventrículos.
Actividad tónica de la inervación autonómica cardíaca
Tanto el simpático como el parasimpático se encuentran siempre activos ejerciendo una acción tónica permanente sobre el corazón, de manera que las funciones cardíacas en reposo son el resultado de la actividad espontánea cardíaca modulada tónicamente por esas dos influencias antagónicas.
El tono parasimpático es mayor que el simpático, lo cual se deduce del hecho de que al “denervar” quirúrgica o farmacológicamente el corazón, este se acelera aumentando la frecuencia cardíaca.
Las demandas metabólicas incrementadas del organismo requieren de un incremento de la actividad cardíaca que se logra automáticamente incrementando la acción que el simpático ejerce sobre el corazón y reduciendo la acción parasimpática. El grado de máximo reposo se logra con las acciones contrarias.
La modulación de los centros cardioacelerador y cardioinhibidor, orígenes mencionados de la inervación autonómica cardíaca, depende de la actividad de centros nerviosos superiores situados en el tallo cerebral, el hipotálamo y la corteza cerebral.