Homocigotos en procariotas y eucariotas
Un homocigoto en genética es un individuo que posee dos copias del mismo alelo (la misma versión de un gen) en uno o más loci (lugar en el cromosoma). El término es aplicado algunas veces a entidades genéticas más grandes como cromosomas completos; en ese contexto, un homocigoto es un individuo con dos copias idénticas del mismo cromosoma.
La palabra homocigoto se compone de dos elementos etimológicamente. Los términos son homo -igual o idéntico- y cigoto -óvulo fertilizado o la primera célula de un individuo originado a través de reproducción sexual-.
Índice del artículo
- 1 Clasificación celular: procariontes y eucariontes
- 2 Homocigotos y dominancia
- 3 Mutaciones dominantes y recesivas
- 4 Homocigotos y herencia
- 5 Genética de poblaciones y evolución
- 6 Referencias
Los organismos son clasificados en función de varias propiedades asociadas al material genético (ADN) contenido en sus células. Considerando la estructura celular donde se localiza el material genético se ha clasificado a los organismos en dos grandes tipos: procariontes (pro: antes; karyon: núcleo) y eucariontes (eu: verdadero; karyon: núcleo).
En los organismos procariontes el material genético está confinado a una región particular en el citoplasma de las células llamada nucleoide. Los organismos modelo de este grupo corresponden a bacterias de la especie Escherichia coli, las cuales presentan una sola cadena de ADN circular, es decir, que sus extremos están unidos entre sí.
A esta cadena se le conoce como cromosoma y en E. coli contiene alrededor de 1.3 millones de pares de bases. Existen algunas excepciones a este patrón dentro del grupo, por ejemplo, algunos géneros bacterianos presentan cromosomas con cadena lineal como las espiroquetas del género Borrelia.
El tamaño lineal o longitud de los genomas/cromosomas bacterianos está generalmente en el rango de milímetros, es decir que son varias veces mayores que el tamaño de las células mismas.
El material genético es guardado en forma empaquetada para reducir el espacio ocupado por esta gran molécula. Este empaquetamiento se logra a través del súper-enrollamiento, un giro sobre el eje principal de la molécula que produce pequeños enroscamientos que al girar causan.
A su vez, enroscamientos mayores de éstas pequeñas roscas sobre sí mismas y el resto de la cadena, reduciendo así la distancia y el espacio ocupado entre distintas secciones del cromosoma circular y llevándolo a una forma condensada (plegado).
En eucariontes el material genético está localizado dentro de un compartimiento especializado rodeado por una membrana; dicho compartimento se conoce como núcleo de la célula.
El material genético contenido dentro del núcleo está estructurado bajo un principio similar al de los procariontes, el súper-enrollamiento.
Sin embargo, los grados/niveles de enroscamiento son mayores dado que la cantidad de ADN a acomodar es mucho mayor. En eucariontes el núcleo no contiene una sola cadena de ADN o cromosoma, contiene varios de ellos y éstos no son circulares, sino lineales y deben ser acomodados.
Cada cromosoma varía en tamaño dependiendo de la especie pero suelen ser más grandes que los de procariontes si se les compara individualmente.
Por ejemplo, el cromosoma 1 de humanos tiene una longitud de 7.3 centímetros, mientras que el cromosoma de E. coli mide 1.6 milímetros aproximadamente. Para mayor referencia, el genoma humano contiene 6.6×109 nucleótidos.
Existe otra clasificación de organismos en función de la cantidad de material genético que contienen, conocida como ploidía.
Los organismos con un solo juego o copia de cromosomas son conocidos como haploides (bacterias o células reproductivas en humanos), con dos juegos/copias de cromosomas son conocidos como diploides (Homo sapiens, Mus musculus, entre muchos otros), con cuatro juegos/copias de cromosomas son conocidos como tetraploides (Odontophrinus americanus, plantas del género Brassicca).
Los organismos con números grandes de juegos de cromosomas son conocidos en conjunto como poliploides. En muchos casos los juegos extra de cromosomas son copias de un conjunto básico.
Durante varios años se consideró que características como la ploidía mayor a uno eran propias de los organismos con núcleo celular definido, pero hallazgos recientes han demostrado que algunos procariontes poseen múltiples copias cromosómicas elevando su ploidía, como demuestran los casos de Deinococcus radiodurans y Bacillus meagateriium.
En organismos diploides (como los chícharos estudiados por Mendel) los dos genes de un loci, o alelos, son heredados uno por vía materna y otro por vía paterna y el par de alelos en conjunto representa el genotipo de ese gen específico.
Un individuo que presenta un genotipo homocigo (homocigoto) para un gen es aquel que posee dos variantes o alelos idénticos en un locus dado.
Los homocigos, a su vez, pueden ser sub-clasificados en dos tipos en función de su relación y contribución al fenotipo: dominantes y recesivos. Cabe resaltar que ambas expresiones son propiedades fenotípicas.
La dominancia en el contexto genético es una relación entre los alelos de un gen en la cual la contribución fenotípica de un alelo es enmascarada por la contribución del otro alelo del mismo locus; en este caso el primer alelo es recesivo y el segundo es dominante (heterocigosis).
La dominancia no es heredada en los alelos o en el fenotipo que producen, es una relación que se establece en función de los alelos presentes y puede ser modificada por agentes externos como otros alelos.
Un ejemplo clásico de dominancia y su relación con el fenotipo es la producción de una proteína funcional por parte del alelo dominante que finalmente produce el rasgo físico, mientras que el alelo recesivo no produce dicha proteína en forma funcional (mutante) y por lo tanto no contribuye al fenotipo.
Así pues, un individuo homocigo dominante para un rasgo/característica es aquel que posee un genotipo que presenta dos copias idénticas del alelo dominante (línea pura).
También es posible encontrar dominancia en genotipos en donde no se encuentran los dos alelos dominantes, sino que se presenta un alelo dominante y uno es recesivo, pero esto no es un caso de homocigosis, es un caso de heterocigosis.
En análisis genéticos, a los alelos dominantes se les representa con una letra mayúscula relacionada al rasgo que se describe.
En el caso de los pétalos de la flor del chícharo, el rasgo silvestre (en este caso el color púrpura) es dominante y el genotipo se representa como “P/P”, denotando tanto al rasgo dominante como a la condición homociga, es decir, la presencia de dos alelos idénticos en un organismo diploide.
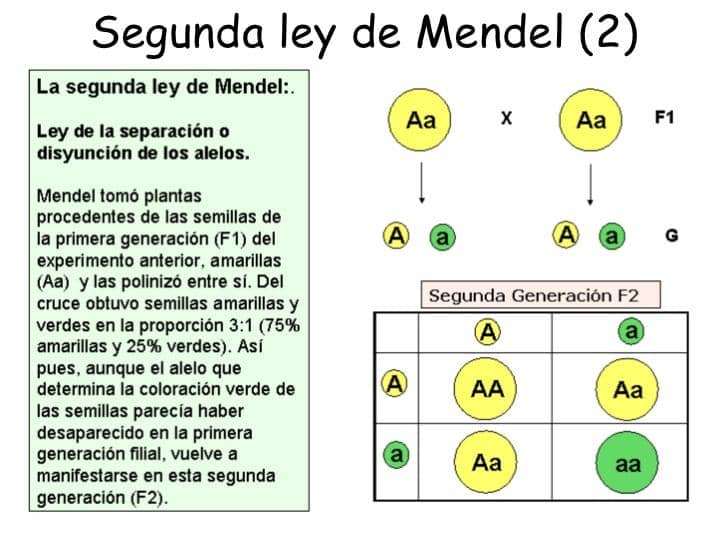
Por su parte, un individuo homocigo recesivo para un rasgo particular porta dos copias del alelo que codifica para el rasgo recesivo.
Siguiendo con el ejemplo del chícharo, el rasgo recesivo en los pétalos es el color blanco, por lo que en individuos con flores de este color cada alelo se representa con una letra minúscula implicando la recesividad y las dos copias idénticas recesivas, por lo que el genotipo es simbolizado como “p/p”.
En algunos casos los genetistas usan de manera simbólica una letra mayúscula para representar al alelo silvestre (por ejemplo P) y con ello simbolizan y hacen referencia a una secuencia nucleotídica específica.
En cambio, cuando se emplea una letra minúscula, p, se representa a un alelo recesivo que puede ser cualquiera de los tipos (mutaciones) posibles[1,4,9].
Los procesos por los cuales un genotipo particular es capaz de producir un fenotipo en los organismos son variados y complejos. Las mutaciones recesivas en general inactivan al gen afectado y llevan a una pérdida de función.
Esto puede pasar por una remoción parcial o completa del gen, por la interrupción de la expresión del gen o por la alteración de la estructura de la proteína codificada que finalmente altera su función.
En cambio, las mutaciones dominantes a menudo producen una ganancia de función, pueden incrementar la actividad de un producto génico dado o conferir una nueva actividad a dicho producto, por lo que también pueden producir una expresión espacio-temporal inapropiada.
Éste tipo de mutaciones también pueden estar asociadas con pérdida de función, existen algunos casos donde dos copias de un gen son requeridas para una función normal de manera que la remoción de una sola copia puede llevar a un fenotipo mutante.
A estos genes se les conoce como haplo-insuficientes. En algunos otros casos la mutación puede llevar a cambios estructurales en las proteínas que interfieren con la función de la proteína silvestre codificada por el otro alelo. A éstas se les conoce como mutaciones dominantes negativas .
En humanos, ejemplos de fenotipos recesivos conocidos son el albinismo, la fibrosis quística y la fenilcetonuria. Todas éstas son condiciones médicas con bases genéticas similares.
Tomando como ejemplo a la última, los individuos con esta enfermedad presentan un genotipo “p/p”, y dado que el individuo posee ambos alelos recesivos, es un homocigoto.
En este caso la “p” está relacionada al término en inglés phenylketonuria y es minúscula para representar el carácter recesivo del alelo. La enfermedad es causada por un procesamiento anormal de la fenilalanina que en condiciones normales debería ser convertida en tirosina (ambas moléculas son aminoácidos) por la enzima fenilalanina hidroxilasa.
Mutaciones en la proximidad del sitio activo de ésta enzima impiden que sea capaz de unir a la fenilalanina para posteriormente procesarla.
Como consecuencia la fenilalanina se acumula en el cuerpo y es convertida en ácido fenilpirúvico, un compuesto que interfiere con el desarrollo del sistema nervioso. Estas condiciones son conocidas en conjunto como desórdenes autosómicos recesivos.
Los patrones de herencia y por tanto la presencia de alelos para un gen, tanto dominantes como recesivos, en los genotipos de los individuos dentro de una población obedecen a la primera ley de Mendel.
Esta ley es conocida como ley de segregación igualitaria de los alelos y tiene bases moleculares que se explican durante la formación de los gametos.
En los organismos diploides que se reproducen sexualmente, existen dos tipos celulares principales: las células somáticas y las células sexuales o gametos.
Las células somáticas poseen dos copias de cada cromosoma (diploides) y cada uno de los cromosomas (cromátidas) contiene a uno de los dos alelos.
Las células gaméticas son producidas por los tejidos germinales a través de meiosis en donde células diploides sufren una división nuclear acompañada de una reducción cromosómica durante dicho proceso, en consecuencia sólo presentan un juego de cromosomas, por lo que son haploides.
Durante la meiosis el huso acromático se ancla a los centrómeros de los cromosomas y las cromátidas son separadas (y por tanto los alelos también) hacia polos opuestos de la célula madre, produciendo dos células hijas separadas o gametos.
Si el individuo productor de los gametos es homocigo (A/A ó a/a) entonces el total de las células gaméticas producidas por él portarán alelos idénticos (A ó a, respectivamente).
Si el individuo es heterocigo (A/a ó a/A) entonces la mitad de los gametos portarán un alelo (A) y la otra mitad el otro (a). Cuando la reproducción sexual se completa se forma un nuevo cigoto, los gametos masculino y femenino se fusionan para formar una nueva célula diploide y se establece una nueva pareja de cromosomas y por tanto de alelos.
Este proceso origina un nuevo genotipo que es determinado por los alelos aportados por el gameto masculino y el gameto femenino.
En genética mendeliana, fenotipos homocigotos y heterocigotos no tienen las mismas probabilidades de aparecer en una población, sin embargo, las posibles combinaciones alélicas asociadas a los fenotipos pueden ser inferidas o determinadas a través de análisis de cruzas genéticas.
Si ambos padres son homocigotos para un gen del tipo dominante (A/A), entonces los gametos de ambos serán del tipo A en su totalidad y su unión resultará en un genotipo A/A invariablemente.
Si ambos padres presentan genotipo homocigo recesivo (a/a), entonces la descendencia invariablemente resultará en un genotipo homocigo recesivo también.
En teoría evolutiva se dice que el motor de la evolución es el cambio y a nivel genético el cambio se da a través de mutaciones y recombinaciones.
Las mutaciones a menudo involucran cambios en alguna base nucleotídica de un gen, aunque pueden ser de más de una base.
A la mayoría de las mutaciones se les considera eventos espontáneos asociados a la tasa de error o fidelidad de las polimerasas durante la transcripción y replicación del ADN.
Existe también mucha evidencia de fenómenos físicos que causan mutaciones a nivel genético. Por su parte, las recombinaciones pueden llegar a producir intercambios de secciones enteras de cromosomas pero sólo están asociadas a eventos de duplicación celular, como la mitosis y la meiosis.
De hecho, se consideran un mecanismo básico para generar variabilidad genotípica durante la formación de gametos. La incorporación de variabilidad genética es una característica distintiva de la reproducción sexual.
Enfocados en los genes, actualmente se considera que el motor de la herencia y por consiguiente la evolución, son los genes que presentan más de un alelo.
Aquellos genes que sólo presentan un alelo difícilmente pueden causar un cambio evolutivo si todos los individuos de la población presentan dos copias del mismo alelo como se ejemplifica arriba.
Esto se debe a que al pasar la información genética de una generación a otra, difícilmente se encontrarán cambios en dicha población a menos que existan fuerzas que produzcan variaciones en los genes como las mencionadas arriba también.
Los modelos evolutivos más sencillos son aquellos que sólo consideran a un locus y su objetivo es tratar de predecir las frecuencias genotípicas en la siguiente generación, a partir de los datos de la generación existente.
- Ridley, M. (2004). Evolutionary Genetics. In Evolution (pp. 95-222). Blackwell Science Ltd.
- Griswold, A. (2008) Genome packaging in prokaryotes: the circular chromosome of E. coli. Nature Education 1(1):57
- Dickerson R.E., Drew H.R., Conner B.N., Wing R.M., Fratini A.V., Kopka, M.L. The anatomy of A-, B-, and Z-DNA. 1982. Science, 216:475-485.
- Iwasa, J., Marshall, W. (2016). Control of Gene Expression. In Karp’s Cell and Molecular Biology, Concepts And Experiments. 8th Edition, Wiley.
- Hartl D.L., Jones E. W. (2005). Genetics: Analysis of Genes and Genomes. pp 854. Jones & Bartlett Learning.
- Mendell, J.E., Clements, K.D., Choat J. H., Angert, E.R.Extreme polyploidy in a large bacterium. 2008. PNAS 105 (18) 6730-6734.
- Lobo, I. & Shaw, K. (2008) Thomas Hunt Morgan, genetic recombination, and gene mapping. Nature Education 1(1):205
- O’Connor, C. (2008) Chromosome segregation in mitosis: The role of centromeres. Nature Education 1(1):28
- Griffiths A.J.F., Wessler, S.R., Lewontin, R.C., Gelbart, W.M., Suzuki, D.T., Miller, J.H. (2005). An Introduction to Genetic Analysis. (pp. 706). W.H. Freeman and Company.
- Lodish, H. F. (2013). Molecular cell biology. New York: W.H. Freeman and Co.