Eritrosa: características, estructura, funciones
La eritrosa es un monosacárido, poseedor de cuatro carbonos, cuya fórmula empírica es C4H8O4. Hay dos azúcares de cuatro carbonos (tetrosas) que derivan del gliceraldehído: la eritrosa y la treosa, siendo ambas polihidroxi-aldehídos (aldosas). La eritrulosa es la única tetrosa que es una polihidroxi-cetona (cetosa). Se deriva de la dihidroxiacetona.
De las tres tetrosas (eritrosa, treosa, eritrulosa) la más común es la eritrosa, que se encuentra en vías metabólicas tales como la vía de la pentosa fosfato, el ciclo de Calvin o las vías de biosíntesis de aminoácidos esenciales y aromáticos.
Índice del artículo
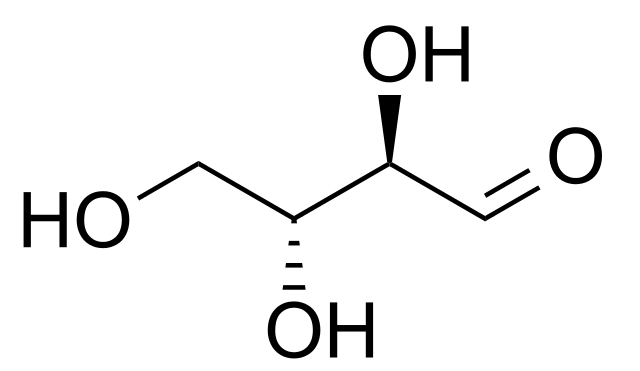
El átomo de carbono uno (C-1) de la eritrosa es el carbono carbonilo de un grupo aldehído (-CHO). Los átomos de carbonos 2 y 3 (C-2 y C-3) son dos grupos hidroximetileno (-CHOH), que son alcoholes secundarios. El átomo de carbono 4 (C-4) es un alcohol primario (-CH2OH).
Los azúcares con la configuración D, tales como la eritrosa, son más abundantes que los azúcares con la configuración L. La eritrosa posee dos carbonos quirales C-2 y C-3, los cuales son centros asimétricos.
En la proyección de Fisher de la eritrosa, el carbono asimétrico más alejado con respecto al grupo carbonilo del aldehído tiene la configuración del D-gliceraldehído. Por lo tanto, el grupo hidroxilo (-OH) del C-3 se representa a la derecha.
La D-eritrosa se diferencia de la D-treosa en la configuración en torno al carbono asimétrico C-2: en la representación de Fisher, el grupo hidroxilo (-OH) de la D-eritrosa se encuentra a la derecha. Por el contrario, en la D-treosa se encuentra a la izquierda.
La adición de un grupo hidroximetileno a la D-eritrosa crea un nuevo centro quiral. Se forman dos azúcares de cinco carbonos (pentosas) de configuración D, a saber: D-ribosa y D-arabinosa, las cuales se diferencian en la configuración del C-2.
En las células, la eritrosa se encuentra en la forma de eritrosa 4-fosfato y se produce a partir de otros azúcares fosforilados. La fosforilación de los azúcares tiene como función elevar su potencial de energía de hidrólisis (o variación de energía de Gibbs, ΔG).
La función química que es fosforilada en los azúcares es el alcohol primario (-CH2OH). Los carbonos de la eritrosa 4-fosfato provienen de la glucosa.
Durante la glicólisis (o ruptura de la molécula de glucosa para obtener energía), el grupo hidroxilo primario del C-6 de la glucosa es fosforilado mediante la transferencia de un grupo fosfato desde el adenosín trifosfato (ATP). Esta reacción es catalizada por la enzima hexoquinasa.
Por otra parte, la síntesis química de azúcares cortos, tales como la D-eritrosa, tiene lugar mediante la oxidación del periodato de 4,6-0-etilideno-O-glucosa, la cual es seguida por la hidrólisis del anillo del acetal.
Alternativamente, aunque no se puede realizar en solución acuosa, se puede usar tetraacetato, el cual corta los a-dioles y además es más estereoespecífico que el ión periodato. La O-glucosa se oxida en presencia de ácido acético, formando 2,3 di-O-formil-D-eritrosa, cuya hidrólisis produce D-eritrosa.
Exceptuando la eritrosa, los monosacáridos se encuentran en su forma cíclica cuando están cristalizados o en solución.
La eritrosa 4-fosfato tiene un papel importante en las vías metabólicas siguientes: vía de la pentosa fosfato, ciclo de Calvin y vías de biosíntesis de aminoácidos esenciales y aromáticos. A continuación se describe la función de la eritrosa 4-fosfato en cada una de estas vías.
La vía de la pentosa fosfato tiene como finalidad producir NADPH, que es el poder reductor de las células, y ribosa 5-fosfato, necesario para la biosíntesis de ácidos nucleicos mediante reacciones oxidativas. El metabolito de partida de esta vía es la glucosa 6-fosfato.
El exceso de ribosa 5-fosfato es convertido en intermediarios glicolíticos. Para ello, son necesarios dos pasos reversibles: 1) reacciones de isomerización y epimerización; 2) reacciones de corte y formación de enlaces C-C que transforman a las pentosas, xilulosa 5-fosfato y ribosa 5-fosfato, en fructosa 6-fosfato (F6P) y gliceraldehído 3-fosfato (GAP).
El segundo paso es llevado a cabo por transaldolasas y transcetolasas. La transaldolasa cataliza la transferencia de tres átomos de carbono (unidad C3) desde la sedoheptulosa 7-fosfato al GAP, produciendo eritrosa 4-fosfato (E4P).
La transcetolasa cataliza la transferencia de dos átomos de carbono (unidad C2) desde la xilulosa 5-fosfato al E4P y forma GAP y F6P.
En el transcurso de la fotosíntesis, la luz proporciona la energía necesaria para la biosíntesis de ATP y NADPH. Las reacciones de fijación del carbono emplean el ATP y el NADPH para reducir el dióxido de carbono (CO2) y formar triosas fosfato mediante el ciclo de Calvin. Luego, las triosas formadas en el ciclo de Calvin son trasformadas en sacarosa y almidón.
El ciclo de Calvin se divide en las tres etapas siguientes: 1) fijación del CO2 en 3-fosfoglicerato; 2) transformación del 3-fosfoglicerato en GAP; y 3) regeneración de la ribulosa 1,5-bifosfato a partir de triosas fosfato.
En la tercera etapa del ciclo de Calvin se forma E4P. Una transcetolasa que contiene pirofosfato de tiamina (TPP) y requiere Mg+2, catalizando la trasferencia de una unidad C2 desde la F6P al GAP, y formando la pentosa xilulosa 5-fosfato(Xu5P) y la tetrosa E4P.
Una aldolasa combina, mediante condensación aldólica, la Xu5P y la E4P para formar la heptosa sedoheptulosa 1,7-bifosfato. Luego siguen dos reacciones enzimáticas que finalmente producen triosas y pentosas.
La eritrosa 4-fosfato y el fosfoenolpiruvato son los precursores metabólicos para la biosíntesis del triptófano, la fenilalanina y la tirosina. En las plantas y bacterias, primero tiene lugar la biosíntesis de corismato, el cual es un intermediario en la biosíntesis de aminoácidos aromáticos.
La biosíntesis de corismato tiene lugar mediante siete reacciones, todas ellas catalizadas por enzimas. Por ejemplo, el paso 6 es catalizado por la enzima 5-enolpiruvilshikimato-3-fosfato, que es inhibida competitivamente por el glifosato (–COO-CH2-NH-CH2-PO3-2). Este último es el ingrediente activo del polémico herbicida RoundUp de Bayer-Monsanto.
El corismato es el precursor de la biosíntesis de triptófano mediante una vía metabólica que implica seis pasos catalizados por enzimas. A través de otra vía, el corismato sirve para la biosíntesis de tirosina y fenilalanina.
- Belitz, H. D., Grosch, W., Schieberle, P. 2009. Food Chemistry, Springer, Nueva York.
- Collins, P.M. 1995. Monosaccharides. Their Chemistry and Their Roles in Natural Products. John Wiley and Sons. Chichester.
- Miesfeld, R. L., McEvoy, M. M. 2017. Biochemistry. W. W. Norton, Nueva York.
- Nelson, D. L., Cox, M. M. 2017. Lehninger principles of biochemistry. W. H. Freeman, Nueva York.
- Voet, D., Voet, J. G., Pratt, C. W. 2008. Fundamentals of biochemistry: life at the molecular level. Wiley, Hoboken.