Fibras de Purkinje: concepto, histología y función
¿Qué son las fibras de Purkinje?
Las fibras de Purkinje cardíacas representan la última etapa del sistema que produce, de manera automática y repetida, la excitación eléctrica requerida para la actividad mecánica ventricular. Se centra en dirigir la excitación hasta los miocitos ventriculares para que estos produzcan la sístole (contracción).
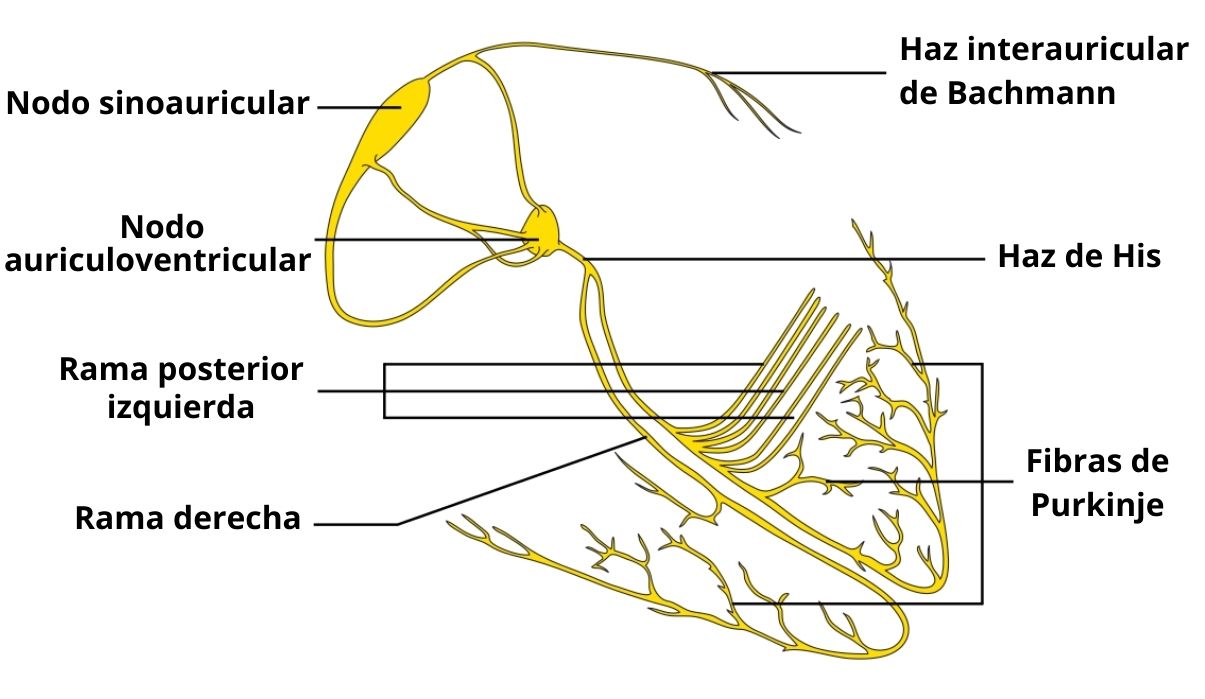
El sistema al que pertenecen estas fibras está integrado por el nodo sino-auricular (SA), en donde se origina la excitación; los fascículos internodales que llegan al nodo aurículo-ventricular (AV); el nodo aurículo-ventricular, en el cual se retrasa un poco la conducción eléctrica; el haz de His, con sus ramas derecha e izquierda, y el sistema de fibras de Purkinje.
Dichas fibras fueron llamadas así en honor de Juan Evangelista Purkinje, anatomista y fisiólogo checo que las describió por primera vez en 1839. No deben ser confundidas con las células de Purkinje, descubiertas por el mismo autor a nivel de la corteza cerebelosa e implicadas en el control del movimiento.
Histología
Al igual que el resto de los componentes del sistema de excitación-conducción cardíacos, las células que conforman el sistema de fibras de Purkinje son células musculares o miocitos cardíacos que han perdido su estructura contráctil y se han especializado en la conducción de la excitación eléctrica.
Sus componentes unen los extremos de las ramificaciones del haz de His y el inicio de una secuencia de miocitos ventriculares, segmentos entre los cuales conducen la excitación eléctrica originada en el nodo sino-auricular, formando una red difusa distribuida por el endocardio que recubre los ventrículos.
Poseen características que las diferencian de los otros componentes del sistema: son fibras más largas y gruesas (40 μm) incluso que las fibras contráctiles ventriculares y tienen la más alta velocidad de conducción: 4 m/s; en comparación con los 1,5 m/s de las que les siguen, las fibras del haz de His.
Esta alta velocidad de conducción se debe, aparte de a su gran diámetro, a que, en sus sitios de contacto, los discos intercalares, existe una gran densidad de uniones en brecha (gap junctions) que permiten el paso fácil de corrientes iónicas entre ellas y la transmisión rápida de la excitación.
Debido a esta alta velocidad de conducción y a la distribución difusa de las fibras de Purkinje, la excitación alcanza de manera casi simultánea al miocardio contráctil de ambos ventrículos, requiriéndose tan solo 0,03 s (30 ms) para completarse la activación de todo el miocardio ventricular.
Función de las fibras de Purkinje
Propiedades eléctricas
Las células del sistema de Purkinje son células excitables que muestran, en reposo, una diferencia de potencial de -90 a -95 mV entre ambas caras de la membrana que separa su interior del líquido extracelular circundante, siendo su interior negativo con respecto al exterior.
Al ser excitadas, estas células responden con una despolarización conocida como potencial de acción (PA) y durante el cual el potencial de la membrana se hace rápidamente menos negativo y puede llegar a invertirse, alcanzando momentáneamente un valor positivo de hasta +30 mV (positivo por dentro).
De acuerdo a la velocidad con la que se produzca esta despolarización, los diferentes tipos celulares excitables del corazón han sido incluidos en una de dos categorías: fibras de respuesta rápida o fibras de respuesta lenta. Las fibras de Purkinje forman parte de esta última categoría.
Potencial de acción en las fibras de Purkinje
El estímulo fisiológico para que las fibras de Purkinje produzcan un potencial de acción es una corriente iónica despolarizante, proveniente de elementos celulares que están antes en la secuencia de conducción, y que les llega a través de las uniones en brecha que las unen con esos elementos.
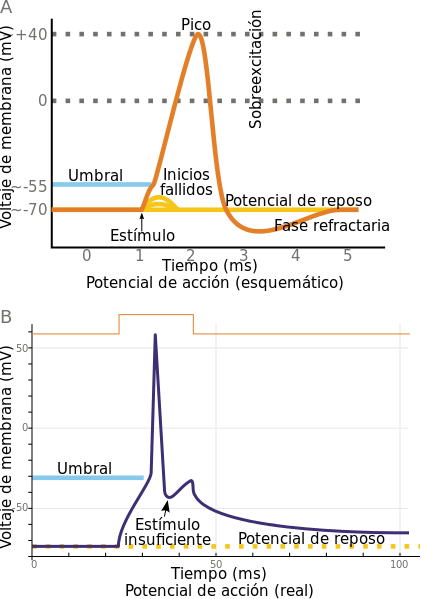
En el potencial de acción de una fibra de Purkinje se distinguen varias fases: una despolarización abrupta (fase 0) hasta +30 mV, una rápida repolarización hasta 0 mV (fase 1), una despolarización sostenida alrededor de los 0 mV (fase 2 o meseta) y una rápida repolarización (fase 3) que lleva de nuevo al potencial de reposo (fase 4).
Estos eventos son el resultado de la activación y/o desactivación de corrientes iónicas que modifican el balance de cargas entre el interior y el exterior de las células. Corrientes que, a su vez, resultan de los cambios de permeabilidad de canales específicos para distintos iones y que se designan con la letra I, seguida de un subíndice que las identifica.
Las corrientes de entrada de iones positivos o salida de iones negativos se consideran por convención negativas y producen despolarizaciones, las de salida de iones positivos o entrada de negativos son corrientes positivas y favorecen la polarización o negativización interna de la célula.
Fases del potencial de acción de las fibras de Purkinje
- La fase 0 se produce cuando la despolarización inicial que sirve de estímulo lleva el potencial de membrana a un nivel (umbral) entre -75 y -65 mV, y se abren entonces unos canales de sodio (Na+) voltaje-dependientes que dejan entrar Na+ (corriente Ina) como en una avalancha, llevando el potencial hasta unos +30 mV.
- La fase 1 comienza al final de la fase 0, cuando los canales de Na+ vuelven a cerrarse y la despolarización se detiene, produciéndose entonces unas corrientes transitorias (Ito1 e Ito2) de salida de K+ y entrada de Cl-, que producen una repolarización rápida hasta el nivel de 0 mV.
- La fase 2 es una “meseta” de larga duración (300 ms). Resulta de la apertura de unos canales de calcio lentos y la producción de una corriente de entrada de Ca++ que mantiene, junto con una entrada remanente de Na+, el potencial relativamente alto (0 mV) y contrarresta las corrientes repolarizantes de K+ (IKr e IKs) que han empezado a producirse.
- En la fase 3 se reducen al mínimo las corrientes de Ca++ y Na+ y las corrientes repolarizantes de salida de K+ se hacen muy pronunciadas. Esta salida cada vez mayor de K+ lleva el potencial de membrana hasta el nivel inicial de reposo de entre -90 y -95 mV en el cual permanece (fase 4) hasta que se repita el ciclo de nuevo.
Valores de algunas propiedades eléctricas de las fibras de Purkinje
- Nivel de reposo: -90 a -95 mV.
- Nivel máximo de despolarización (overshoot): + 30 mV.
- Amplitud del potencial de acción: 120 mV.
- Duración del potencial de acción: entre 300 y 500 ms.
- Velocidad de la despolarización: 500-700 V/s.
- Nivel umbral para el disparo del potencial de acción: entre -75 y -65 mV.
- Velocidad de conducción: 3-4 m/s.
Fibras de Purkinje como marcapasos secundarios
Las fibras miocárdicas de respuesta lenta incluyen a las células de los nodos sino-auricular y aurículo-ventricular, las cuales, durante el reposo (fase 4), van sufriendo una despolarización lenta (prepotencial diastólico) que lleva el potencial de membrana hasta su nivel umbral y se dispara de manera automática un potencial de acción.
Esta propiedad está más desarrollada, es decir, la despolarización ocurre más rápidamente, en el nodo sino-auricular, que sirve como marcapaso cardíaco y marca una frecuencia de entre 60 y 80 lat/min. Si falla, el nodo aurículo-ventricular puede asumir el comando, pero con una frecuencia más baja de entre 60 y 40 lat/min.
Las fibras de Purkinje, cuando no reciben excitación por la vía del sistema de conducción normal, pueden sufrir también ese mismo proceso de despolarización lenta que lleva su potencial de membrana hasta el nivel umbral, y terminar disparando potenciales de acción de manera automática.
En caso de que falle la excitación normal del nodo sino-auricular y la secundaria del nodo aurículo-ventricular, o se bloquee el paso de la excitación a los ventrículos, algunas fibras del sistema de Purkinje empiezan a descargar por su cuenta y mantienen una activación ventricular rítmica, pero a una frecuencia más baja (25-40 lat/min).