Beta oxidación de ácidos grasos: pasos, reacciones, productos, regulación
La beta oxidación de ácidos grasos es la ruta del catabolismo (degradación) de los ácidos grasos, que tiene como función principal la producción o la “liberación” de la energía contenida en los enlaces de estas moléculas.
Esta ruta fue descubierta en 1904 gracias a los experimentos realizados por el alemán Franz Knoop, que consistieron en la administración, a ratas experimentales, de ácidos grasos cuyo grupo metilo final había sido modificado con un grupo fenilo.
Knoop esperaba que los productos del catabolismo de estos ácidos grasos “análogos” siguieran rutas similares a la ruta de oxidación de los ácidos grasos normales (los naturales no modificados). Sin embargo, encontró que existían diferencias en los productos obtenidos en función del número de átomos de carbono de los ácidos grasos.
Con dichos resultados, Knoop propuso que la degradación ocurría en “escalones”, comenzando con un “ataque” al carbono β (el de la posición 3 respecto al grupo carboxilo terminal), liberando unos fragmentos de dos átomos de carbono.
Posteriormente fue demostrado que el proceso requiere energía en forma de ATP, que se produce en las mitocondrias y que los fragmentos de dos átomos de carbono ingresan al ciclo de Krebs como acetil-CoA.
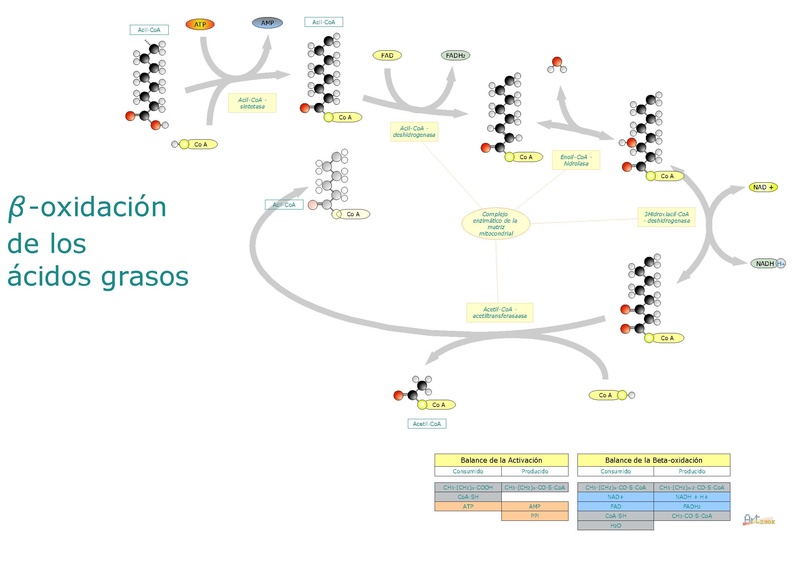
En resumidas cuentas, la beta oxidación de ácidos grasos implica la activación del grupo carboxilo terminal, el transporte del ácido graso activado hacia la matriz mitocondrial y la oxidación “escalonada” de dos en dos carbonos desde el grupo carboxilo.
Como muchos procesos anabólicos y catabólicos, esta ruta es regulada, puesto que amerita de la movilización de los ácidos grasos de “reserva” cuando las otras rutas catabólicas no son suficientes para cubrir las demandas energéticas celulares y corporales.
Índice del artículo
- 1 Pasos y reacciones
- 1.1 – Activación de ácidos grasos y transporte hacia la mitocondria
- 1.2 – Beta oxidación de ácidos grasos saturados con un número par de átomos de carbono
- 1.3 – Beta oxidación de ácidos grasos saturados con un número impar de átomos de carbono
- 1.4 – Beta oxidación de ácidos grasos insaturados
- 1.5 – Beta oxidación extramitocondrial
- 2 Productos de la beta oxidación
- 3 Regulación
- 4 Referencias
Los ácidos grasos están predominantemente en el citosol, ya provengan de las rutas biosintéticas o de los depósitos de grasas que son almacenadas a partir de los alimentos ingeridos (que deben ingresar a las células).
La activación de los ácidos grasos requiere la utilización de una molécula de ATP y tiene que ver con la formación de conjugados tioésteres de acilo con la coenzima A.
Esta activación es catalizada por un grupo de enzimas llamadas acetil-CoA ligasas específicas respecto a la longitud de la cadena de cada ácido graso. Algunas de estas enzimas activan los ácidos grasos a medida que son transportados hacia la matriz mitocondrial, pues están embebidas en la membrana mitocondrial externa.
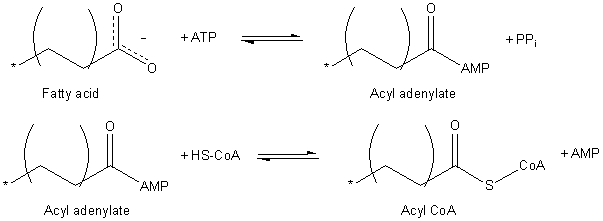
El proceso de activación ocurre en dos pasos, produciéndose primero un acil adenilato a partir del ácido graso activado con el ATP, donde se libera una molécula de pirofosfato (PPi). El grupo carboxilo activado por el ATP es después atacado por el grupo tiol de la coenzima A formando acil-CoA.
La translocación del acil-CoA a través de la membrana mitocondria se consigue gracias a un sistema de transporte conocido como la lanzadera de carnitina.
La degradación de los ácidos grasos es una ruta cíclica, puesto que la liberación de cada fragmento de dos átomos de carbono es seguida inmediatamente por otra, hasta alcanzar la longitud total de la molécula. Las reacciones que tienen parte en este proceso son las siguiente:
– Deshidrogenación.
– Hidratación de un doble enlace.
– Deshidrogenación de un grupo hidroxilo.
– Fragmentación por el ataque de una molécula de acetil-CoA sobre el carbono β.
Reacción 1: primera deshidrogenación
Consiste en la formación de un doble enlace entre el carbono α y el carbono β por eliminación de dos átomos de hidrógeno. Es catalizada por una enzima acil-CoA deshidrogenasa, que forma una molécula de trans∆2-enoil-S-CoA y una molécula de FAD+ (cofactor).
Reacciones 2 y 3: hidratación y deshidrogenación
La hidratación es catalizada por la enoil-CoA hidratasa, entretanto la deshidrogenación es mediada por la 3-hidroxiacil-CoA deshidrogenasa, y esta última reacción depende del cofactor NAD+.
La hidratación del trans∆2-enoil-S-CoA da origen a un 3-hidroxiacil-CoA, cuya deshidrogenación produce una molécula de 3-cetoacil-CoA y un NADH + H.
El FADH2 y el NADH producidos en las primeras tres reacciones de la beta oxidación son reoxidados a través de la cadena transportadora de electrones, gracias a lo cual participan de la producción de ATP, 2 moléculas por cada FADH2 y 3 moléculas por cada NADH.
Reacción 4: fragmentación
Cada ciclo de beta oxidación que elimina una molécula de dos átomos de carbono finaliza con la fragmentación “tiolítica” del carbono ceto, el cual es atacado por la coenzima A en el enlace entre los carbonos α y β.
Esta reacción es catalizada por la enzima β-cetotiolasa o tiolasa, y sus productos son una molécula de acil-CoA (el ácido graso activado con dos átomos de carbono menos) y una de acetil-CoA.
En aquellos ácidos grasos de número impar de átomos de carbono (que no son muy abundantes), la molécula del último ciclo de degradación tiene 5 átomos de carbono, por lo que su fragmentación produce una molécula de acetil-CoA (que ingresa al ciclo de Krebs) y otra de propionil-CoA.
El propionil-CoA debe ser carboxilado (reacción dependiente de ATP y bicarbonato) por la enzima propionil-CoA carboxilasa, con lo que se forma un compuesto conocido como D-metilmalonil-CoA, que debe ser epimerizado a su forma “L”.
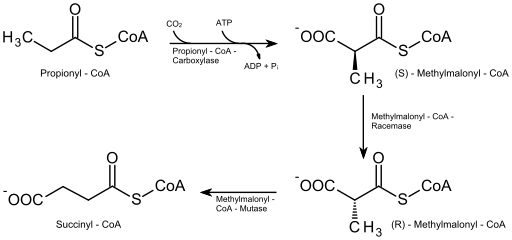
El compuesto resultante de la epimerización es convertido después en succinil-CoA por acción de la enzima L-metilmalonil-CoA mutasa, y esta molécula, así como el acetil-CoA, ingresa al ciclo de los ácidos cítricos.
Muchos lípidos celulares tienen cadenas de ácidos grasos con insaturaciones, es decir, que poseen uno o más enlaces dobles entre sus átomos de carbono.
La oxidación de estos ácidos grasos es un poco diferente a la de los ácidos grasos saturados, pues dos enzimas adicionales, la enoil-CoA isomerasa y la 2,4-dienoil-CoA reductasa, se encargan de eliminar dichas insaturaciones para que estos ácidos grasos puedan ser sustrato de la enzima enoil-CoA hidratasa.
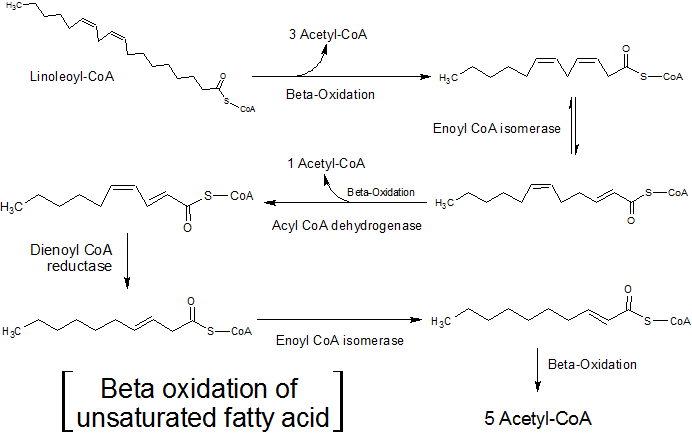
La enoil-CoA isomerasa actúa sobre ácidos grasos monoinsaturados (con una sola insaturación), entretanto la enzima 2,4-dienoil-CoA reductasa reacciona con los ácidos grasos poliinsaturados (con dos o más insaturaciones).
La beta oxidación de los ácidos grasos también puede darse en el interior de otros orgánulos citosólicos como los peroxisomas, por ejemplo, con la diferencia de que los electrones que son transferidos al FAD+ no son entregados a la cadena respiratoria, sino directamente al oxígeno.
Esta reacción produce peróxido de hidrógeno (el oxígeno es reducido), compuesto que es eliminado por la enzima catalasa, específica de estos orgánulos.
La oxidación de ácidos grasos produce mucha mayor cantidad de energía que la degradación de carbohidratos. El principal producto de la beta oxidación es el acetil-CoA producido en cada paso de la porción cíclica de la ruta, sin embargo, otros productos son:
– AMP, H+ y pirofosfato (PPi), producidos durante la activación.
– FADH2 y NADH, por cada acetil-CoA producido.
– Succinil-CoA, ADP, Pi, para los ácidos grasos de cadena impar.
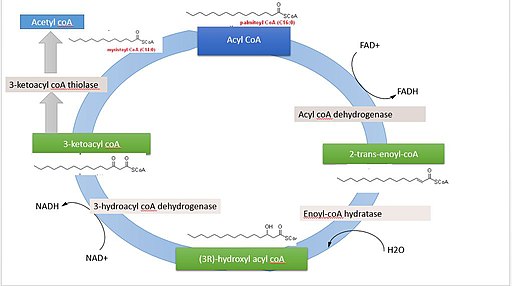
Si consideramos como ejemplo la beta oxidación completa del ácido palmítico (palmitato) un ácido graso de 16 átomos de carbonos, la cantidad de energía que se produce es equivalente más o menos a 129 moléculas de ATP, que provienen de las 7 vueltas que debe completar el ciclo.
La regulación de la beta oxidación de ácidos grasos en la mayor parte de las células depende de la disponibilidad energética, no solo relacionada con los carbohidratos sino con los mismos ácidos grasos.
Los animales controlan la movilización y, por ende, la degradación de las grasas mediante estímulos hormonales, que al mismo tiempo son controlados por moléculas como el AMPc, por ejemplo.
En el hígado, el principal órgano de degradación de grasas, la concentración de malonil-CoA es sumamente importante para la regulación de la beta oxidación; este es el primer sustrato comprometido en la ruta de biosíntesis de los ácidos grasos.
Cuando el malonil-CoA se acumula en grandes proporciones promueve la biosíntesis de ácidos grasos e inhibe el transportador mitocondrial o la lanzadera acil-carnitina. Cuando su concentración disminuye, la inhibición cesa y se activa la beta oxidación.
- Mathews, C., van Holde, K., & Ahern, K. (2000). Biochemistry (3th ed.). San Francisco, California: Pearson.
- Nelson, D. L., & Cox, M. M. (2009). Lehninger Principios de Bioquímica. Ediciones Omega (5th ed.).
- Rawn, J. D. (1998). Biochemistry. Burlington, Massachusetts: Neil Patterson Publishers.
- Schulz, H. (1991). Beta oxidation of fatty acids. Biochimica et Biophysica Acta, 1081, 109–120.
- Schulz, H. (1994). Regulation of Fatty Acid Oxidation in Heart. Critical Review, 165–171.
- Schulz, H., & Kunau, W. (1987). Beta-oxidation of unsaturated fatty acids : a revised pathway. TIBS, 403–406.