Opsoninas: funciones, tipos, receptores
Las opsoninas son moléculas del sistema inmune que se unen al antígeno y a las células inmunes conocidas como fagocitos, facilitando el proceso de fagocitosis. Algunos ejemplos de células fagocíticas que pueden participar en este proceso son los macrófagos.
Una vez que un patógeno supera las barreras anatómicas y fisiológicas del hospedador, es posible que cause infección y enfermedad. Por ello, el sistema inmunitario reacciona a esta invasión detectando al cuerpo extraño por medio de sensores y atacándolo con un elaborado mecanismo de respuesta.
Índice del artículo
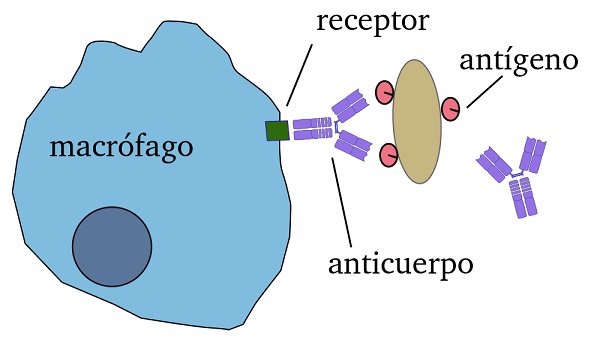
Las opsoninas recubren las partículas a fagocitar mediante la interacción con los antigenos. De esta manera, las células fagocíticas como mácrofagos y células dendríticas, que expresan receptores para opsoninas, se unen a los patógenos opsonisados mediante estos receptores y finalmente los fagocitan.
De modo que, las opsoninas actúan como una especie de puente entre el fagocito y la partícula a ser fagocitada.
Las opsoninas se encargan de contrarrestar la fuerza repelente entre las paredes celulares negativas y promueven la absorción del patógeno por parte del macrófago.
Sin la acción de opsoninas, las paredes celulares cargadas negativamente del patógeno y el fagocito se repelen entre sí, entonces el agente extraño puede burlar su destrucción y continuar replicándose dentro del hospedador.
Entonces, la opsonización es una estrategia antimicrobiana para frenar y eliminar la propagación de una enfermedad.
Existen diversos tipos de opsoninas, entre ellas la lectina de unión a manosa, las inmunoglobulinas del isotipo IgG y componentes del sistema de complemento como C3b, iC3b o C4b.
La lectina de unión a manosa es producida en el hígado y es liberada a la sangre. Tiene la capacidad de unirse a repeticiones de azúcares presentes en los microorganismos favoreciendo su destrucción mediante la activación del sistema de complemento a través de la asociación de serín proteasas.
IgG es el único isotipo de inmunoglobulina que tiene la capacidad de atravesar la placenta, debido a su pequeño tamaño. Existen 4 subisotipos, los cuales presentan funciones específicas.
C3b, es el principal componente formado tras la ruptura de la proteína C3 del sistema de complemento.
iC3b, es formado cunado el factor I del complemento escinde a la proteína C3b.
Finalmente, C4b es el producto de la proteólisis del C1q, el cual es un complejo de proteínas que ante la formación de complejos antígeno-anticuerpo se activan siguiendo una secuencia.
Es importante destacar que la opsonización de un patógeno puede ocurrir por medio de anticuerpos o del sistema del complemento.
Los anticuerpos son parte del sistema inmunitario adaptativo, que son producidos por las células plasmáticas en respuesta a un determinado antígeno. Un anticuerpo tiene una compleja estructura que le confiere especificidad a ciertos antígenos.
Al final de las cadenas pesada y ligera, los anticuerpos tienen regiones variables (sitios de unión a antígeno), que le permiten al anticuerpo encajar como “una llave en un candado”. Una vez que los sitios de unión al antígeno son ocupados, la región del tallo del anticuerpo se une al receptor en los fagocitos.
De este modo, el patógeno queda envuelto por el fagosoma y es destruido por los lisosomas.
Además, el complejo antígeno-anticuerpo también puede activar el sistema de complemento. La inmunoglobulina M (IgM), por ejemplo, es muy eficiente en la activación del complemento.
Los anticuerpos IgG también son capaces de unirse a las células efectoras inmunes a través de su dominio constante, desencadenando una liberación de productos de lisis de la célula efectora inmunitaria.
El sistema de complemento por su parte, posee más de 30 proteínas que mejoran la capacidad de los anticuerpos y las células fagocíticas para combatir los organismos invasores.
Las proteínas del complemento, identificadas con la letra “C” de complemento, están formadas por 9 proteínas (C1 a C9), las cuales están inactivas cuando circulan por todo el cuerpo humano. Sin embargo, cuando un patógeno es detectado, las proteasas escinden los precursores inactivos y los activan.
Ahora bien, la respuesta del organismo a la presencia de un patógeno o cuerpo extraño puede llevarse a cabo a través de tres vías: la clásica, la alternativa y la vía de las lectinas.
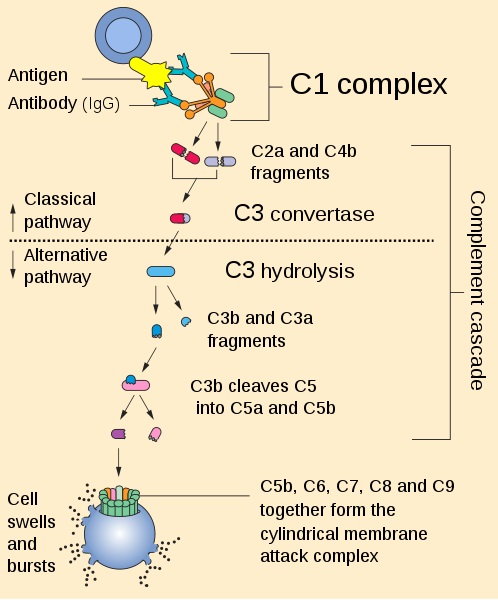
El MAC, está constituido por un complejo de proteínas del complemento, las cuales se asocian a la parte externa de la membrana plasmática de bacterias patógenas y forman una especie de poro. El objetivo final de la formación del poro es provocar la lisis del microorganismo.
Una vez que se ha generado C3b, por cualquiera de las vías del sistema del complemento, Este se une a múltiples sitios en la superficie celular del patógeno y luego se agrega a los receptores expresados en la superficie del macrófago o neutrófilo.
En los leucocitos se expresan cuatro tipos de receptores que reconocen fragmentos de C3b: CR1, CR2, CR3 y CR4. La deficiencia en estos receptores hace a la persona más susceptible a sufrir de continuas infecciones.
C4b, al igual que C3b, puede unirse al receptor CR1. Mientras que iC3b se une al CR2.
Entre los receptores Fc destacan los FcℽR, que reconocen distintos subisotipos de IgG.
La unión de la partícula opsonizada a los receptores de fagocitos de la superficie celular (receptores Fc), desencadena la formación de pseudópodos que rodean la partícula extraña de manera similar a una cremallera a través de interacciones receptor-opsonina.
Cuando los pseudópodos se encuentran, se fusionan formando una vacuola o fagosoma, que luego se une con el lisosoma en el fagocito, el cual descarga una batería de enzimas y especies de oxígeno tóxico antibacteriano, iniciando la digestión de la partícula extraña para eliminarla.
- McCulloch J, Martin SJ. Assays of cellular activity. 1994. Cellular Immunology, pp.95-113.
- Roos A, Xu W, Castellano G, Nauta AJ, Garred P, Daha MR, van Kooten C. Mini-review: A pivotal role for innate immunity in the clearance of apoptotic cells. European Journal of Immunology. 2004; 34(4): 921-929.
- Sarma JV, Ward PA. The complement system. Cell and tissue research. 2011; 343(1), 227-235.
- Thau L, Mahajan K. Physiology, Opsonization. 2018. StatPearls Publishing. Recuperado de https://www.ncbi.nlm.nih.gov/books/NBK534215/
- Thomas J, Kindt Richard A. Goldsby Amherst College Barbara A. Osborne. Javier de León Fraga (Ed.). 2006. En Inmunología de Kuby sexta edición. pp. 37, 94-95.
- Wah S, Aimanianda V. Host Soluble Mediators: Defying the Immunological Inertness of Aspergillus fumigatus Conidia. Journal of Fungi. 2018; 4(3): 1-9.
- Zhang Y, Hoppe AD, Swanson JA. Coordination of Fc receptor signaling regulates cellular commitment to phagocytosis. Proceedings of the National Academy of Sciences. 2010; 107(45):19332-9337.