Hemolisina: características, tipos, mecanismos de acción
La hemolisina es una pequeña proteína que causa poros en la membrana celular de los eritrocitos y algunas otras células propias de la sangre de los mamíferos. Generalmente es sintetizada y excretada por bacterias patógenas.
Esta proteína es una de las toxinas microbianas más comunes y la que mejor se ha estudiado. En ocasiones puede causar anemia hemolítica, ya que la cantidad de canales a través de los que sale el interior celular puede incluso causar la lisis celular.

Generalmente, la hemolisina es una toxina típica de las especies de Streptococcus del tracto intestinal. Su función permite a las bacterias romper la barrera epitelial del tracto intestinal y así desplazarse a través del torrente sanguíneo hasta colonizar otros tejidos.
La forma más común en que se encuentra la hemolisina en la naturaleza es en su forma α-hemolisina. Esta proteína es uno de los factores de virulencia más importantes de la mayoría de las cepas de Escherichia coli y algunos clostridios.
La mayoría de las infecciones del tracto urinario son causadas por cepas de Escherichia coli que producen α-hemolisina con características hemolíticas.
Se ha relacionado la producción de hemolisina y bacteriocina en las cepas bacterianas con un mecanismo de competencia contra las otras especies y la producción de ambas toxinas parece depender de los mismos determinantes genéticos en el genoma de las bacterias.
Índice del artículo
La hemolisina está constituida por siete subunidades y el gen que la codifica posee siete promotores. Estas siete subunidades se insertan en la membrana plasmática de las células blanco y, al juntarse, forman un canal iónico por donde se escapan los metabolitos del interior celular.
La hemolisina es una citotoxina dependiente de calcio (Ca+2) extracelular que actúa sobre la membrana plasmática de las células del torrente sanguíneo. Los poros que crea en la membrana también son hidrofílicos y causan la entrada de agua en el interior celular, lo que puede ocasionar la lisis.
Las hemolisinas son productos proteicos típicos de las bacterias de tipo gram-negativas y todas comparten dos características:
1- La presencia de un péptido muy pequeño (nonapéptido) conformado por repetidos de los aminoácidos glicina y ácido aspártico. Los nonapéptidos de la hemolisina se ubican cerca de la porción C-terminal de la estructura primaria de la proteína.
2- Todas las hemolisinas son secretadas por la bacteria al medio extracelular a través de un transportador de tipo ABC (del inglés ATP-Binding Cassette).
Habitualmente se detecta la producción de hemolisinas en las cepas bacterianas a través de un crecimiento en medio agar sangre. En la prueba, se observa un halo hemolítico, producto de la ruptura de los glóbulos rojos cerca de las colonias bacterias.
Existen varios tipos de hemolisinas diferentes, estas se clasifican con una letra griega al comienzo de su nombre. Las más estudiadas y comunes son las α, las β y las γ hemolisinas, todas producidas por la cepa Staphylococcus aureus.
Los tipos de hemolisina se clasifican según el rango de células que atacan y según su la estructura primaria de la proteína.
Esta proteína es típica de las cepas de Staphylococcus aureus y Escherichia coli; ataca a neutrófilos, glóbulos rojos, linfocitos, macrófagos, fibroblastos adultos y embrionarios. Interactúa con las cabezas polares de los lípidos de membrana plasmática de estas células hasta internalizar una cola hidrofóbica de cerca de 5 Ӑ en el interior de la membrana.
Producida por Staphylococcus aureus en menor proporción que la α-hemolisina, la β-hemolisina ataca principalmente a los eritrocitos y se interna en la membrana exclusivamente a través de los dominios ricos en esfingomielina de la membrana celular.
También se ha observado en Staphylococcus aureus. Ha sido catalogada como una proteína hemolítica y leucotoxina al mismo tiempo, ya que afecta a las células polimorfonucleares de humanos, monocitos, macrófagos y rara vez, incluso a los glóbulos rojos.
Este tipo de γ-hemolisina es una de las que menos se ha caracterizado, por tanto, se desconoce gran parte de su mecanismo de acción y este no se ha investigado in vivo.
El mecanismo de acción que se ha esclarecido de forma relativamente clara es el de la α-hemolisina. Sin embargo, ya que todas son proteínas hemolíticas, se piensa que la mayoría de los procesos son comunes para todas las hemolisinas.
Los científicos sugieren que para las bacterias segreguen hemolisina al ambiente tienen que encontrarse en un microambiente pobre en nutrientes, por tanto, este sería un mecanismo que dispara la célula para destruir a las células blanco y obtener sus nutrientes.
El mecanismo se ha descrito en tres pasos: la unión a la membrana celular, la inserción y la oligomerización.
Se ha encontrado que las hemolisinas pueden unirse a las integrinas de los neutrófilos y en los eritrocitos se ha observado que estas proteínas se unen a componentes glicosilados como las glicoproteínas, los gangliósidos y las glicoforinas de la membrana celular.
Algunos autores sugieren que la presencia de receptores en la membrana no es esencial para que ocurra la unión de las hemolisinas. En todo caso, aún no se conoce con precisión el mecanismo de reconcomiendo celular de la proteína.
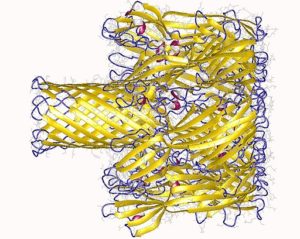
La interacción con la membrana ocurre en dos pasos:
– Unión inicial (reversible): cuando la hemolisina se une a los dominios de unión a calcio de la membrana. Este paso ocurre en la superficie y es muy susceptible a descargas electroestáticas.
– Unión irreversible: junta los dominios aminoacídicos con los componentes lipídicos de la capa externa de la membrana plasmática de las células blanco, con el fin de formar uniones físicas entre los compuestos hidrofóbicos de la membrana.
La α-hemolisina inserta los residuos 177 y 411 en la primera monocapa lipídica. En el medio extracelular la hemolisina se asocia con iones de calcio, los cuales inducen un arreglo estructural en esta y contribuyen con su activación.
Esta inserción consolida la unión irreversible a la membrana de la célula. Una vez ha ocurrido el arreglo, la hemolisina se transforma en una proteína integral puesto que, experimentalmente, se ha demostrado que la única forma de extraerla de la membrana es mediante el uso de detergentes como el Tritón X-100.
Cuando toda la hemolisina se ha insertado a la membrana plasmática de las células blanco tiene lugar la oligomerización de las 7 subunidades que la conforman, lo que termina en la formación de un poro proteico, muy dinámico pero dependiente de la composición lipídica de la membrana.
Se ha observado que el proceso de oligomerización es favorecido por los microdominios o las balsas lipídicas de la membrana celular. Estas regiones quizás no favorecen la unión de la proteína, pero si favorecen la oligomerización de la misma una vez insertada.
Entre mayor cantidad de hemolisinas se unen a la membrana, mayor cantidad de poros se formarán. Es más, las hemolisinas pueden oligomerizarse entre sí (las adyacentes) y conformar canales mucho más grandes.
- Bakás, L., Ostolaza, H., Vaz, W. L., & Goñi, F. M. (1996). Reversible adsorption and nonreversible insertion of Escherichia coli alpha-hemolysin into lipid bilayers. Biophysical journal, 71(4), 1869-1876.
- Dalla Serra, M., Coraiola, M., Viero, G., Comai, M., Potrich, C., Ferreras, M., … & Prévost, G. (2005). Staphylococcus aureus bicomponent γ-hemolysins, HlgA, HlgB, and HlgC, can form mixed pores containing all components. Journal of chemical information and modeling, 45(6), 1539-1545.
- Gow, J. A., & Robinson, J. (1969). Properties of Purified Staphylococcal β-Hemolysin. Journal of bacteriology, 97(3), 1026-1032.
- Ike, Y., Hashimoto, H., & Clewell, D. B. (1984). Hemolysin of Streptococcus faecalis subspecies zymogenes contributes to virulence in mice. Infection and Immunity, 45(2), 528-530.
- Remington, J. S., Klein, J. O., Wilson, C. B., Nizet, V., & Maldonado, Y. A. (Eds.). (1976). Infectious diseases of the fetus and newborn infant (Vol. 4). Philadelphia: Saunders.
- Todd, E. W. (1932). Antigenic streptococcal hemolysin. Journal of experimental medicine, 55(2), 267-280.