Operón lac: descubrimiento y función
El operón lac es un grupo de genes estructurales que tiene como función codificar para proteínas involucradas en el metabolismo de la lactosa. Son unos genes que están ordenados de forma consecutiva en el genoma de casi todas las bacterias y se han estudiado con especial esfuerzo en la bacteria “modelo” Escherichia coli.
El operón lac fue que el modelo empleado por Jacob y Monod en 1961 para la proposición de los arreglos genéticos en forma de operón. En sus trabajos, dichos autores describieron cómo la expresión de uno o más genes podía “encenderse” o “apagarse” como consecuencia de la presencia de una molécula (la lactosa, por ejemplo) en el medio de crecimiento.
Las bacterias que crecen en medios de crecimiento ricos en compuestos carbonados o azúcares diferentes a la lactosa, como la glucosa y la galactosa, poseen cantidades muy bajas de las proteínas necesarias para la metabolización de la lactosa.
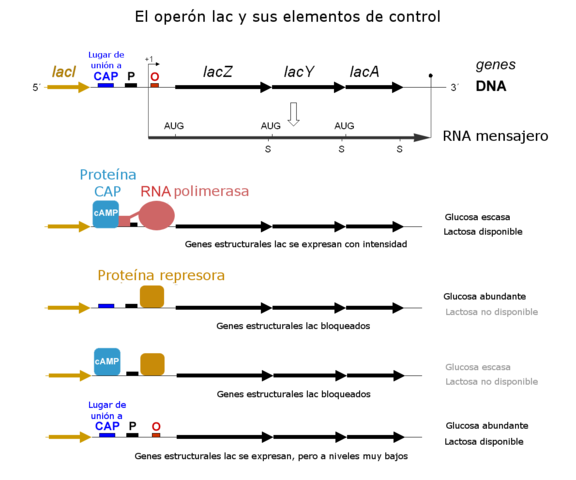
Entonces, en ausencia de lactosa el operón se encuentra “apagado”, evitando que la ARN polimerasa transcriba al segmento génico correspondiente al operón lac. Cuando la célula “percibe” la presencia de lactosa se activa el operón y estos genes se transcriben normalmente, lo que se conoce como “encendido” del operón.
Todos los genes del operón se traducen en una sola molécula de ARN mensajero y, por lo tanto, cualquier factor que regule la transcripción de este ARN mensajero del operón lac regulará directamente la transcripción de cualquier gen que pertenezca al mismo.
Índice del artículo
La teoría de Jacob y Monod se desarrolló en un contexto donde se sabía muy poco sobre la estructura del ADN. Y es que tan solo ocho años antes Watson y Crick habían realizado su propuesta sobre la estructura del ADN y del ARN, por lo que los ARN mensajeros apenas se conocían.
Jacob y Monod en la década de 1950 ya habían demostrado que el metabolismo de lactosa bacteriano estaba genéticamente regulado por dos condiciones muy específicas: la presencia y la ausencia de lactosa.
Ambos científicos habían observado que una proteína con características similares a una enzima alostérica era capaz de detectar la presencia de lactosa en el medio y, que una vez detectado el azúcar, se estimula la transcripción de dos enzimas: una lactosa permeasa y otra galactosidasa.
Hoy en día se sabe que la permeasa ejerce funciones en el transporte de la lactosa hacia el interior de la célula y que la galactosidasa es necesaria para “romper” o “cortar” la molécula de lactosa en glucosa y galactosa, de modo que la célula pueda aprovechar este disacárido en sus partes constituyentes.
Cerca de la década de 1960 ya se había determinado que la lactosa permeasa y la galactosidasa eran codificadas por dos secuencias genéticas adyacentes, la región Z y la región Y, respectivamente.
Finalmente, en 1961, Jacob y Monod expusieron un modelo genético compuesto por cinco elementos genéticos:
– Un promotor
– Un operador y
– los genes Z, Y y A.
Todos estos segmentos se traducen en un único ARN mensajero y comprenden las partes esenciales para definir prácticamente a cualquier operón bacteriano en la naturaleza.
Jacob, Monod y sus colaboradores realizaron muchos experimentos con células bacterianas que poseían mutaciones que hacían a las cepas incapaces de metabolizar lactosa. Tales cepas se identificaban con el nombre de la cepa y la mutación correspondiente que poseían.
De esta manera los investigadores fueron capaces de identificar que las mutaciones en los genes lacZ, que codifica para la β-galactosidasa, y lacY, que codifica para la lactosa permeasa, producían bacterias del tipo lac–, es decir, bacterias incapaces de metabolizar lactosa.
A partir de “mapeos genéticos” utilizando enzimas de restricción se determinó posteriormente la ubicación de los genes en las diferentes cepas, hecho que permitió establecer que los tres genes lacZ, lacY y lacA se encuentran (en ese orden) en el cromosoma bacteriano en un grupo de genes adyacentes.
La existencia de otra proteína, denominada proteína represora, que no necesariamente se considera como “parte” del operón, fue dilucidada a través de mutaciones de un gen llamado lacI-. Este codifica para una proteína que se une a la región del “operador” en el operón y evita la transcripción de los genes para la β-galactosidasa y la lactosa permeasa.
Se dice que esta proteína no forma parte de los genes que componen al operón lac, pues se ubican, en realidad, “aguas arriba” de estos últimos y son transcritos en ARN mensajeros diferentes.
Las cepas bacterianas que poseen la mutación lacI- expresan “constitutivamente” los genes lacZ, lacY y lacA, lo que se da independientemente de la presencia o ausencia de lactosa en el entorno extracelular.
Muchas de estas observaciones fueron corroboradas mediante la transferencia de los genes lacI+ y lacZ+ a una célula bacteriana que no producía las proteínas codificadas por estos genes en un medio carente de lactosa.
Puesto que las bacterias “transformadas” de esta manera solo producían la enzima β-galactosidasa en presencia de lactosa, el experimento confirmó que el gen lacI era importante para la regulación de la expresión del operón lac.
El operón lac regula la transcripción de los genes que son necesarios para que las bacterias asimilen la lactosa como fuente de carbono y energía. Sin embargo, la trascripción de estos genes solo ocurre cuando la fuente de energía principal corresponde a los carbohidratos de tipo galactósido.
En las células bacterianas existen mecanismos que regulan la expresión de los genes del operón lac cuando estas están en presencia de glucosa o de cualquier otro azúcar más “fácil” de metabolizar.
La metabolización de estos azúcares implica su transporte al interior celular y su ruptura o procesamiento posterior.
La lactosa es utilizada como una fuente de energía alterna para las bacterias, ayudándoles a sobrevivir incluso después de que se agotan otras fuentes de energía del medio como la glucosa.
El modelo del operón lac fue el primer sistema genético de este tipo en ser dilucidado y, por lo tanto, sirvió como base para describir muchos otros operones en el genoma de distintos tipos de microorganismos.
Con el estudio de este sistema se avanzó ampliamente en el conocimiento del funcionamiento de las proteínas de tipo “represor” que se unen al ADN. También se avanzó en el entendimiento de las enzimas alostéricas y de cómo actúan selectivamente al reconocer uno u otro sustrato.
Otro importante avance que surgió a partir del estudio del operón lac fue el establecimiento de la función crucial que juegan los ARNs mensajeros para traducir las instrucciones que se encuentran en el ADN y también como paso previo a la síntesis proteica.
- Griffiths, A. J., Wessler, S. R., Lewontin, R. C., Gelbart, W. M., Suzuki, D. T., & Miller, J. H. (2005). An introduction to genetic analysis. Macmillan.
- Hartwell, L., Goldberg, M. L., Fischer, J. A., Hood, L. E., & Aquadro, C. F. (2008). Genetics: from genes to genomes (pp. 978-0073227382). New York: McGraw-Hill.
- Lewis, M. (2013). Allostery and the lac Operon. Journal of molecular biology, 425(13), 2309-2316.
- Müller-Hill, B., & Oehler, S. (1996). The lac operon (pp. 66-67). New York:: Walter de Gruyter.
- Parker, J. (2001). lac Operon.
- Yildirim, N., & Kazanci, C. (2011). Deterministic and stochastic simulation and analysis of biochemical reaction networks: The lactose operon example. In Methods in enzymology (Vol. 487, pp. 371-395). Academic Press.