Ley de Hardy-Weinberg: qué es y ejemplo
¿Qué es la ley de Hardy-Weinberg?
La ley deHardy-Weinberg, también llamada principio o equilibrio de Hardy-Weinberg, consiste en un teorema matemático que describe a una población hipotética diploide con reproducción sexual que no está evolucionando, es decir, que las frecuencias alélicas no cambian de generación en generación.
Dicho principio asume cinco condiciones necesarias para que la población se mantenga constante: ausencia de flujo génico, ausencia de mutaciones, apareamiento aleatorio, ausencia de selección natural y un tamaño poblacional infinitamente grande. En ausencia de dichas fuerzas, la población se mantiene en equilibrio.
Cuando alguno de los supuestos anteriores no se cumple, ocurre un cambio. Por esta razón, la selección natural, la mutación, las migraciones y la deriva genética son los cuatro mecanismos evolutivos.
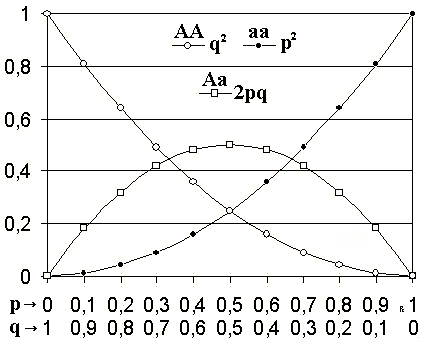
Según este modelo, cuando las frecuencias alélicas de una población son p y q, las frecuencias genotípicas serán p2, 2pq y q2.
Se puede aplicar el equilibrio de Hardy-Weinberg en el cálculo de las frecuencias de ciertos alelos de interés, por ejemplo, para calcular la proporción de heterocigotos en una población humana.
También podemos verificar si una población se encuentra o no en el equilibrio y proponer hipótesis de qué fuerzas están actuando en dicha población.
¿En qué consiste la teoría de Hardy-Weinberg?
La teoría o equilibrio de Hardy-Weinberg es un modelo nulo que nos permite especificar el comportamiento de las frecuencias génicas y alélicas a lo largo de las generaciones.
En otras palabras, es el modelo que describe el comportamiento de los genes en las poblaciones, bajo una serie de condiciones específicas.
Notación
En el teorema de Hardy-Weinberg, la frecuencia alélica de A (alelo dominante) se representa con la letra p, mientras que la frecuencia alélica de a (alelo recesivo) se representa con la letra q.
Las frecuencias genotípicas esperadas son p2, 2pq y q2, para el homocigoto dominante (AA), heterocigoto (Aa) y homocigoto recesivo (aa), respectivamente.
Si solo hay dos alelos en dicho locus, la suma de las frecuencias de los dos alelos necesariamente tiene que ser igual a 1 (p + q = 1).
La expansión de binomio (p + q)2 representan las frecuencias genotípicas p2 + 2pq + q2 = 1.
Ejemplo
En una población, los individuos que la integran se cruzan entre sí para dar origen a la descendencia. De manera general, podemos puntualizar los aspectos más importantes de este ciclo reproductivo: la producción de gametos, la fusión de estos para dar origen a un cigoto, y el desarrollo del embrión para dar lugar a la nueva generación.
Imaginemos que podemos rastrear el proceso de los genes mendelianos en los acontecimientos mencionados. Hacemos esto porque deseamos conocer si un alelo o genotipo aumentará o disminuirá su frecuencia y por qué lo hace.
Para entender cómo varían las frecuencias génicas y alélicas en una población, seguiremos la producción de gametos de un conjunto de ratones.
En nuestro ejemplo hipotético, el apareamiento ocurre al azar, donde todos los espermatozoides y los óvulos se mezclan de manera aleatoria.
En el caso de los ratones, esta suposición no es cierta y es solo una simplificación para facilitar los cálculos. Sin embargo, en algunos grupos animales, como ciertos equinodermos y otros organismos acuáticos, los gametos son expulsados y colisionan al azar.
Primera generación de ratones
Ahora, centremos nuestra atención en un locus concreto, con dos alelos: A y a. Siguiendo la ley enunciada por Gregor Mendel, cada gameto recibe un alelo de locus A. Supongamos que el 60% de los óvulos y del esperma reciben el alelo A, mientras que el 40% restante recibieron el alelo a.
Por esto, la frecuencia del alelo A es 0,6 y la del alelo a es de 0,4. Este grupo de gametos se encontrará al azar para dar origen a un cigoto, ¿qué probabilidad hay para que formen cada uno de los tres genotipos posibles? Para ello, debemos multiplicar las probabilidades de la siguiente manera:
Genotipo AA: 0,6 x 0,6 = 0,36.
Genotipo Aa: 0,6 x 0,4 = 0,24. En el caso del heterocigoto, existen dos forman en las cuales se puede originar. La primera, que el esperma porte el alelo A y el óvulo el alelo a, o el caso inverso, el esperma el a y el óvulo A. Por ello sumamos 0,24 + 0,24 = 0,48.
Genotipo aa: 0,4 x 0,4 = 0,16.
Segunda generación de ratones
Ahora bien, imaginemos que estos cigotos se desarrollan y se convierten en ratones adultos que nuevamente producirán gametos, ¿esperaríamos que las frecuencias alélicas sean iguales o diferentes a la generación anterior?
El genotipo AA producirá el 36% de los gametos, mientras que los heterocigotos producirán el 48% de los gametos, y el genotipo aa el 16%.
Para calcular la nueva frecuencia del alelo, sumamos la frecuencia del homocigoto más la mitad del heterocigoto, de la siguiente manera:
Frecuencia del alelo A: 0,36 + ½ (0,48) = 0,6.
Frecuencia del alelo a: 0,16 + ½ (0,48) = 0,4.
Si las comparamos con las frecuencias iniciales, nos daremos cuenta de que son idénticas. Por ello, según el concepto de evolución, como no hay cambios en las frecuencias alélicas a lo largo de las generaciones, la población está en equilibrio, no evoluciona.
Suposiciones del equilibrio de Hardy-Weinberg
¿Qué condiciones debe cumplir la población anterior para que sus frecuencias alélicas se mantengan constantes con el paso de las generaciones? En el modelo del equilibrio Hardy-Weinberg, la población que no evoluciona cumple con los siguientes supuestos:
La población es infinitamente grande
– La población debe tener un tamaño extremadamente grande para evitar los efectos estocásticos o aleatorios de la deriva génica.
– Cuando las poblaciones son pequeñas, el efecto de la deriva génica (cambios aleatorios en las frecuencias alélicas, de una generación a otra) debido al error de muestreo es mucho mayor y puede producir la fijación o la pérdida de ciertos alelos.
No existe flujo génico
– Las migraciones no existen en la población, por lo que no pueden llegar o salir alelos que puedan alterar las frecuencias génicas.
No hay mutaciones
– Las mutaciones son cambios en la secuencia de ADN, y pueden tener distintas causas. Estos cambios aleatorios modifican el acervo génico en la población, por la introducción o eliminación de genes en los cromosomas.
Apareamiento aleatorio
– La mezcla de los gametos debe realizarse de forma aleatoria –como la suposición que usamos en el ejemplo de los ratones–. Por ello, no debe existir una elección de pareja entre los individuos de la población, incluida la endogamia (reproducción de individuos que están emparentados).
– Cuando el apareamiento no es aleatorio, no causa un cambio en las frecuencias alélicas de una generación a la próxima, pero sí puede generar desviaciones de las frecuencias genotípicas esperadas.
No hay selección
– No existe un éxito reproductivo diferencial de los individuos con distintos genotipos que pueda alterar las frecuencias de los alelos dentro de la población.
En otras palabras, en la población hipotética todos los genotipos tienen la misma probabilidad de reproducirse y de sobrevivir.
Cuando una población no cumple con estas cinco condiciones, el resultado es la evolución. Lógicamente, las poblaciones naturales no cumplen con estos supuestos. Por ello, se usa el modelo de Hardy-Weinberg como hipótesis nula que nos permite hacer estimaciones aproximadas de las frecuencias génicas y alélicas.
Además de la falta de estas cinco condiciones, existen otras posibles causas por las que la población no está en equilibrio.
Una de estas ocurre cuando los loci están ligados al sexo o los fenómenos de distorsión en la segregación o meiotic drive (cuando cada copia de un gen o cromosoma no es transmitida con igual probabilidad a la siguiente generación).
Referencias
- Andrews, C. (2010). The Hardy-Weinberg Principle. Nature Education Knowledge.
- Futuyma, D. J. (2005). Evolution. Sinauer.
- Soler, M. (2002). Evolución: la base de la Biología. Proyecto Sur.