¿Qué es el dihibridismo?
¿Qué es el dihibridismo?
El dihibridismo, en genética, define el estudio simultáneo de dos caracteres hereditarios distintos, y por extensión, de aquellos otros cuya manifestación depende de dos genes diferentes, aunque se trate del mismo carácter.
Los siete rasgos que Gregor Mendel analizó le fueron útiles en el planteamiento de su teoría de la herencia de los caracteres porque, entre otras cosas, los genes responsables de su manifestación poseían alelos contrastantes cuyo fenotipo era fácil de analizar, y porque cada uno determinaba la expresión de un único carácter.
Es decir, eran rasgos monogénicos cuya condición híbrida (monohíbridos) permitía determinar las relaciones de dominancia/recesividad entre los alelos de ese único gen.
Cuando Mendel analizó la herencia conjunta de dos caracteres distintos, procedió como lo había hecho con los caracteres sencillos. Obtuvo híbridos dobles (dihíbridos) que le permitieron comprobar dos cosas:
– Que cada uno cumplía con la segregación independiente que había observado en los cruces monohíbridos.
– Que, además, en los cruces dihíbridos la manifestación de cada carácter era independiente de la manifestación fenotípica del otro. Es decir, que sus factores de herencia, cualesquiera que fueran, se distribuían independientemente.
Ahora sabemos que la herencia de los caracteres es un poco más compleja que lo que observó Mendel, pero también que en sus fundamentos Mendel estaba completamente en lo cierto.
El desarrollo posterior de la genética permitió demostrar que los cruces dihíbridos y su análisis (dihibridismo), tal como pudo demostrarlo inicialmente William Bateson, podían ser una fuente inagotable de descubrimientos en esta poderosa y naciente ciencia del siglo XX.
Gracias a su empleo inteligente, podían dar al genetista una idea más clara del comportamiento y naturaleza de los genes.
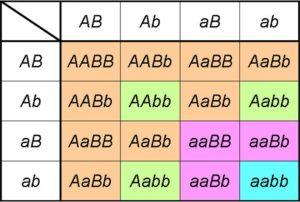
Los cruces dihíbridos de caracteres distintos
Si analizamos los productos de un cruce monohíbrido Aa X Aa, podemos notar que es igual a desarrollar el producto notable (A+a)2= AA + 2Aa + aa.
La expresión de la izquierda incluye los dos tipos de gametos que puede producir uno de los parentales heterocigotos para el gen A/a. Al elevar al cuadrado indicamos que ambos parentales son de idéntica constitución para el gen en estudio [es decir, es un cruce monohíbrido (A+a) X (A+a)].
La expresión a la derecha nos proporciona los genotipos (y, por tanto, se deducen los fenotipos) y proporciones esperadas derivadas del cruce.
De ahí que podamos observar directamente las proporciones genotípicas derivadas de la primera ley (1:2:1), así como las proporciones fenotípicas explicadas por la misma (1AA + 2Aa = 3A_ por cada 1aa, o proporción fenotípica 3:1).
Si consideramos ahora un cruce para analizar la herencia de un gen B, las expresiones y proporciones serán las mismas. De hecho, será así para cualquier gen. En un cruce dihíbrido, por lo tanto, tenemos realmente el desarrollo de los productos de (A+a)2 X (B+b)2.
O lo que es lo mismo, si el cruce dihíbrido implica a dos genes que participan en la herencia de dos caracteres no relacionados entre sí, las proporciones fenotípicas serán aquellas predichas por la segunda ley: (3A_:1aa) X (3B_:1bb)= 9A_B_:3A_bb:3aaB_:1aabb).
Estas, por supuesto, derivan de las proporciones genotípicas ordenadas 4:2:2:2:2:1:1:1:1 que se obtienen como resultado del producto de (A+a)2 X (B+b)2=(AA + 2Aa + aa) X (BB + 2Bb + bb).
Te invitamos a comprobarlo por ti mismo a fin de analizar ahora qué ocurre cuando las proporciones fenotípicas 9:3:3:1 de un cruce dihíbrido se “desvían” de estas claras y predecibles relaciones matemáticas que explican la herencia independiente de dos caracteres codificados por genes distintos.
Manifestaciones fenotípicas alternativas de los cruces dihíbridos
Hay dos maneras principales en que los cruces dihíbridos se alejan de lo “esperado”. La primera es aquella en la cual estamos analizando la herencia conjunta de dos caracteres distintos, pero las proporciones fenotípicas observadas en la progenie dan un claro predominio a la manifestación de los fenotipos parentales.
Lo más probable es que se trate de un caso de genes ligados. Es decir, los dos genes bajo análisis, aunque estén en loci distintos, están tan cercanos el uno del otro físicamente que tienden a heredarse juntos y, obviamente, no se distribuyen independientemente.
La otra circunstancia, que es también bastante común, deriva del hecho de que una exigua minoría de los caracteres hereditarios son monogénicos.
Por el contrario, en la manifestación de la mayoría de los caracteres hereditarios participan más de dos genes.
Por esta razón, cabe siempre la posibilidad de que las interacciones génicas que se establezcan entre los genes que participan en la manifestación de un único carácter, sean complejas y vayan más allá que de una simple relación de dominancia o recesividad, tal como se observa en las relaciones alélicas típicas de los rasgos monogénicos.
Por ejemplo, en la manifestación de un carácter puede que participen unas cuatro enzimas en un orden particular para dar origen al producto final responsable de la manifestación fenotípica del fenotipo silvestre.
El análisis que permite identificar el número de genes de distintos loci que participan en la manifestación de un rasgo genético, así como el orden en el que actúan, se denomina análisis de epistasis, y es tal vez el que define más típicamente lo que denominamos análisis genético en su sentido más clásico.
Referencias
- Brooker, R. J. Genetics: Analysis and Principles. McGraw-Hill Higher Education, New York, USA.
- Cordell, H. Epistasis: What it means, what it doesn’t mean, and statistical methods to detect it in humans. Human Molecular Genetics.