Receptores muscarínicos: estructura, tipos y sus funciones, antagonistas
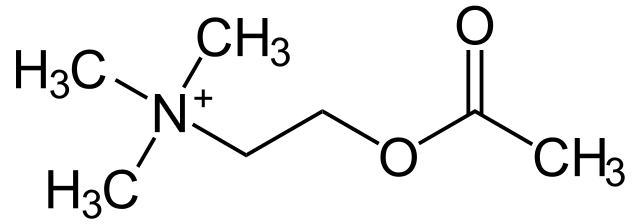
Los receptores muscarínicos son moléculas que median acciones de la acetilcolina (ACh) y se ubican en la membrana postsináptica de las sinapsis en las cuales se libera dicho neurotransmisor; su nombre procede de su sensibilidad al alcaloide muscarina producido por el hongo Amanita muscaria.
En el sistema nervioso central existen varios conjuntos neuronales cuyos axones liberan acetilcolina. Algunos de ellos terminan en el encéfalo mismo, mientras que la mayor parte conforma las vías motoras para el músculo esquelético o las vías efectoras del sistema nervioso autónomo para las glándulas y los músculos cardíaco y liso.
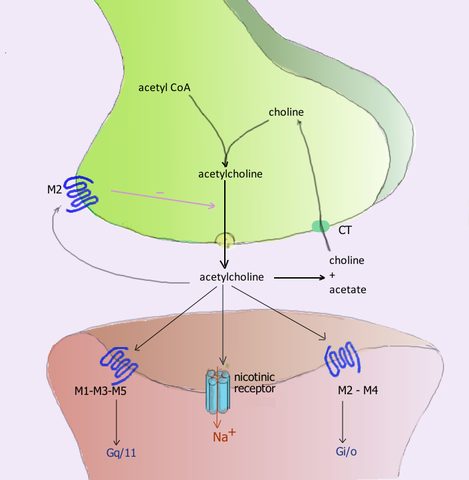
La acetilcolina liberada en las uniones neuromusculares del músculo esquelético activa receptores colinérgicos llamados nicotínicos, por su sensibilidad al alcaloide nicotina, y que también se encuentran en las sinapsis ganglionares del sistema nervioso autónomo (SNA).
Las neuronas postganglionares de la división parasimpática de este sistema ejercen sus funciones liberando acetilcolina, que actúa sobre los receptores colinérgicos muscarínicos ubicados en las membranas de las células efectoras, e induciendo en ellas modificaciones eléctricas por cambios de permeabilidad en sus canales iónicos.
Índice del artículo
Los receptores muscarínicos pertenecen a la familia de los receptores metabotrópicos, término con el cual se designa a aquellos receptores que no son propiamente canales iónicos, sino estructuras proteicas que al activarse desencadenan procesos metabólicos intracelulares que modifican la actividad de los verdaderos canales.
El término se usa para diferenciarlos de los receptores ionotrópicos, los cuales sí son verdaderos canales iónicos que se abren o cierran por acción directa del neurotransmisor, como es el caso de los receptores nicotínicos ya nombrados de las placas neuromusculares del músculo esquelético.
Dentro de los receptores metabotrópicos, los muscarínicos se incluyen en el grupo conocido como receptores acoplados a proteína G, porque dependiendo de su tipo su acción es mediada por algunas variantes de dicha proteína, como Gi, inihibidora de la adenilciclasa, y Gq o G11 que activan a la fosfolipasa C (PLC).
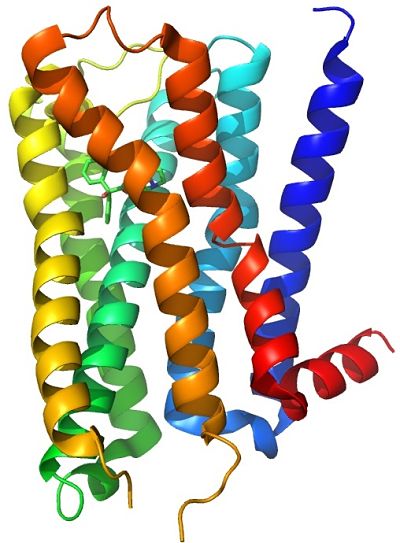
Los receptores muscarínicos son largas proteínas integrales de membrana; tienen siete segmentos transmembranales compuestos por hélices alfa, que cruzan secuencialmente la bicapa lipídica membranal. Por dentro, en la cara citoplásmica, se asocian con la proteína G correspondiente que transduce la interacción ligando-receptor.
Se han identificado al menos 5 tipos de receptores muscarínicos que se designan empleando la letra M seguida de un número, a saber: M1, M2, M3, M4 y M5.
Los receptores M1, M3 y M5 forman la familia M1 y se caracterizan por su asociación a proteínas Gq o G11, mientras que los receptores M2 y M4 son de la familia M2 y se asocian a la proteína Gi.
Se encuentran principalmente en el sistema nervioso central, en las glándulas exocrinas y en los ganglios del sistema nervioso autónomo. Se acoplan a proteína Gq, que activa a la enzima fosfolipasa C, la cual convierte al fosfatidil inositol (PIP2) en inositol trifosfato (IP3), que libera Ca++ intracelular, y diacilglicerol (DAG), que activa a la proteincinasa C.
Se encuentran fundamentalmente en el corazón, principalmente en las células del nodo sinoauricular, sobre las cuales actúan disminuyendo su frecuencia de descarga, como a continuación se describe.
Automatismo cardíaco
Los receptores M2 han sido estudiados más a fondo a nivel del nodo sinoauricular (SA) del corazón, lugar en donde se manifiesta normalmente el automatismo que produce periódicamente las excitaciones rítmicas responsables de la actividad mecánica cardíaca.
Las células del nodo sinoauricular, después de cada potencial de acción (PA) desencadenante de una sístole cardíaca (contracción), se repolarizan y vuelven al nivel de unos -70 mV. Pero el voltaje no permanece en ese valor, sino que sufre una despolarización progresiva hasta un nivel umbral que desencadena un nuevo potencial de acción.
Esta despolarización progresiva se debe a cambios espontáneos en corrientes iónicas (I) que incluyen: reducción de salida de K+ (IK1), aparición de una corriente de entrada de Na+ (If) y luego una entrada de Ca++ (ICaT), hasta que se alcanza el umbral y se dispara otra corriente de Ca++ (ICaL) responsable del potencial de acción.
Si la salida de K+ (IK1) es muy baja y las corrientes de entrada de Na+ (If) y Ca++ (ICaT) son altas, la despolarización ocurre más rápido, el potencial de acción y la contracción se producen más temprano, y la frecuencia cardíaca es más alta. Las modificaciones contrarias en esas corrientes bajan la frecuencia.
Cambios metabotrópicos inducidos por la noradrenalina (simpático) y la acetilcolina (parasimpático) pueden alterar esas corrientes. El AMPc activa directamente los canales de If, la proteincinasa A (PKA) fosforila y activa los canales de Ca++ de ICaT y el grupo βγ de proteína Gi activa la salida de K+.
Acción muscarínica M2
Cuando la acetilcolina liberada por las terminaciones postganglionares de las fibras vagales (parasimpáticas) cardíacas se une a los receptores muscarínicos M2 de las células del nodo sinoauricular, la subunidad αi de la proteína Gi cambia su GDP por GTP y se separa, dejando libre al bloque βγ.
La subunidad αi inhibe a la adenilciclasa y reduce la producción de AMPc, lo cual reduce la actividad de los canales para If y la de la PKA. Este último hecho reduce la fosforilación y actividad de los canales de Ca++ para ICaT; el resultado es una reducción de las corrientes despolarizantes.
El grupo formado por las subunidades βγ de la proteína Gi activa una corriente de K+ hacia afuera (IKACh) que tiende a contrarrestar las entradas de Na+ y Ca++ y baja la velocidad de despolarización.
El resultado conjunto es una reducción de la pendiente de despolarización espontánea y una reducción de la frecuencia cardíaca.
Se pueden encontrar en el músculo liso (aparato digestivo, vejiga, vasos sanguíneos, bronquios), en algunas glándulas exocrinas y en el sistema nervioso central.
Se acoplan también a proteína Gq y, a nivel pulmonar pueden provocar broncoconstricción, mientras que actuando en el endotelio vascular liberan óxido nítrico (NO) y provocan vasodilatación.
Estos receptores están menos caracterizados y estudiados que los anteriores. Se ha reportado su presencia en el sistema nervioso central y en algunos tejidos periféricos, pero sus funciones no están claramente establecidas.
El antagonista universal para estos receptores es la atropina, un alcaloide extraído de la planta Atropa belladona, que se une a ellos con una alta afinidad, lo cual representa un criterio para diferenciarlos de los receptores nicotínicos que son insensibles a esta molécula.
Existe una gran cantidad de otras sustancias antagonistas que se unen a los distintos tipos de receptores muscarínicos con afinidades distintas. La combinación de distintos valores de afinidad por algunas de ellas ha servido precisamente para la inclusión de estos receptores en una u otra de las categorías descritas.
Una lista parcial de otros antagonistas incluiría: pirenzepina, metoctramina, 4-DAMP, himbacina, AF-DX 384, tripitramina, darifenacina, PD 102807, AQ RA 741, pFHHSiD, MT3 y MT7; toxinas estas últimas contenidas en los venenos de las mambas verde y negra, respectivamente.
Los receptores M1, por ejemplo, presentan una elevada sensibilidad por pirenzepina; los M2 por tripitramina, metoctramina e himbacina; los M3 por 4-DAMP; los M4 son muy afines a la toxina MT3 y además a la himbacina; los M5 se parecen mucho a los M3, pero con respecto a ellos son menos afines por AQ RA 741.
- Ganong WF: Neurotransmitters and Neuromodulators, in: Review of Medical Physiology, 25th ed. New York, McGraw-Hill Education, 2016.
- González JC: Papel de los receptors muscarínicos en la modulación de la transmisión GABAérgica en hipocampo. Memoria para optar al grado de doctor. Universidad Autónoma de Madrid. 2013.
- Guyton AC, Hall JE: Rythmical excitation of the Heart, in: Textbook of Medical Physiology , 13th ed; AC Guyton, JE Hall (eds). Philadelphia, Elsevier Inc., 2016.
- Piper HM: Herzerregung, in: Physiologie des Menschen mit Pathophysiologie, 31th ed; RF Schmidt et al (eds). Heidelberg, Springer Medizin Verlag, 2010.
- Schrader J, Gödeche A, Kelm M: Das Hertz, in: Physiologie, 6th ed; R Klinke et al (eds). Stuttgart, Georg Thieme Verlag, 2010.
- Siegelbaum SA, Clapham DE, Schwartz JH: Modulation of Synaptic Transmission: Second Messengers, In: Principles of Neural Science, 5th ed; E Kandel et al (eds). New York, McGraw-Hill, 2013.