¿Qué es el paquiteno y qué ocurre en él?
El paquiteno o paquinema es la tercera etapa de la profase I meiótica; en ella se verifica el proceso de recombinación. En la mitosis hay una profase, y en la meiosis dos: la profase I y la profase II.
Previamente, excepto la profase II, se duplicaron los cromosomas dando origen cada uno a una cromátida hermana. Pero solo en la profase I se aparean los homólogos (duplicados), formando bivalentes.

El término paquiteno proviene del griego y significa “hilos gruesos”. Estos “hilos gruesos” son los cromosomas homólogos apareados que, tras duplicarse, forman tétradas. Es decir, cuatro “hilos”, o cuerdas, que hacen ver a cada cromosoma engrosado.
Existen aspectos únicos de la profase I meiótica que explican las características propias del paquiteno. Solo en el paquiteno de la profase I de la meiosis los cromosomas recombinan.
Para ello se verifica el reconocimiento y apareamiento de los homólogos. Como en la mitosis, debe haber duplicación de las cromátidas. Pero solo en el paquiteno de la meiosis I se forman complejos de intercambio de bandas que llamamos quiasmas.
En ellos ocurre lo que define el poder recombinatorio de la meiosis: el entrecruzamiento entre cromátidas de cromosomas homólogos.
Todo el proceso de intercambio de ADN es posible gracias a la aparición previa del complejo sinaptonémico. Este complejo multiproteico permite que los cromosomas homólogos entren en apareamiento (sinapsis) y recombinen.
Índice del artículo
- 1 El complejo sinaptonémico durante el paquiteno
- 2 Componentes del complejo sinaptonémico y los quiasmas
- 3 Progresión del paquiteno
- 4 Referencias
El complejo sinaptonémico (CS) es el entramado proteico que permite la unión de extremo a extremo entre los cromosomas homólogos. Solo ocurre durante el paquiteno de la meiosis I, y es el fundamento físico del apareamiento cromosomal. En otras palabras, es lo que permite que los cromosomas entren en sinapsis y recombinen.
El complejo sinaptonémico está sumamente conservado entre los eucariotas que experimentan meiosis. Por lo tanto, es evolutivamente muy antiguo, y estructural y funcionalmente equivalente en todos los seres vivos.
Consiste en un elemento axial central y dos elementos laterales que se van repitiendo como los dientes de una cremallera o cierre.
El complejo sinaptonémico se va formado a partir de puntos específicos en los cromosomas durante el zigoteno. Estos sitios son colineares con aquellos donde ocurren las rupturas del ADN donde se experimentará sinapsis y recombinación en el paquiteno.
Durante el paquiteno, por lo tanto, tenemos una cremallera cerrada. En esta conformación terminan de definirse puntos específicos donde se intercambiarán bandas de ADN al final del estadio.
El complejo sinaptonémico meiótico contiene muchas proteínas estructurales que se encuentran también durante la mitosis. Estas incluyen la topoisomerasa II, condensinas, cohesinas, así como proteínas asociadas a las cohesinas.
Además de estas, también están presentes proteínas que son específicas y únicas de la meiosis, junto con proteínas propias del complejo recombinatorio.
Estas proteínas forman parte del recombinosoma. Esta estructura agrupa a todas las proteínas requeridas para la recombinación. Aparentemente el recombinosoma no se forma sobre los puntos de entrecruzamiento, sino que es reclutado, ya formado, hacia ellos.
Los quiasmas son las estructuras morfológicas visibles en los cromosomas donde ocurren los entrecruzamientos. Dicho de otra manera, la manifestación física del intercambio de bandas de ADN entre dos cromosomas homólogos. Los quiasmas son las marcas citomorfológicas distintivas del paquiteno.
En toda meiosis debe ocurrir al menos un quiasma por cromosoma. Esto quiere decir que todo gameto es recombinante. Gracias a este fenómeno se pudieron deducir y proponer los primeros mapas genéticos basados en ligamiento y recombinación.
Por otro lado, la carencia de quiasmas, y por lo tanto de entrecruzamiento, origina distorsiones a nivel de la segregación cromosómica. La recombinación durante el paquiteno actúa entonces como un control de calidad de la segregación meiótica.
Sin embargo, evolutivamente hablando no todos los organismos experimentan recombinación (por ejemplo, los machos de la mosca de la fruta). En estos casos, operan otros mecanismos de segregación cromosómica no dependientes de la recombinación.
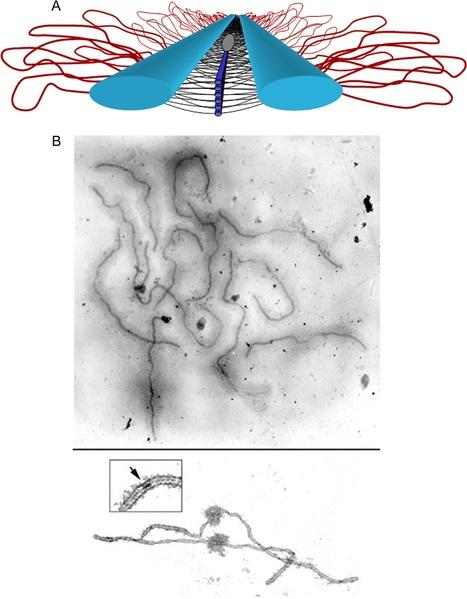
Al salir del zigoteno, el complejo sinaptonémico está completamente formado. Esto se ve complementado con la generación de las rupturas de ADN doble banda a partir de las cuales se verifican los entrecruzamientos.
Las rupturas dobles del ADN obligan a la célula a repararlos. En el proceso de reparación del ADN la célula recluta al recombinosoma. Se recurre al intercambio de bandas, y como resultado, se obtienen células recombinantes.
Cuando está completamente formado el complejo sinaptonémico, se dice que comienza el paquiteno.
Los bivalentes en sinapsis en el paquiteno interactúan básicamente a través del elemento axial del complejo sinaptonémico. Cada cromátida está organizada en una organización de lazos, cuya base es el elemento axial central del complejo sinaptonémico.
El elemento axial de cada homólogo contacta con el del otro a través de los elementos laterales. Los ejes de las cromátidas hermanas están altamente compactados, y sus lazos cromatínicos emergen hacia fuera del elemento axial central. El espaciado entre los lazos (~20 por micrómetro) está evolutivamente conservado entre todas las especies.
Hacia el término del paquiteno, se hacen evidentes entrecruzamientos a partir de algunos de los sitios de ruptura del ADN doble banda. La aparición de los entrecruzamientos señala igualmente el comienzo del desentramado del complejo sinaptonémico.
Los cromosomas homólogos se condensan más (se ven más individuales) y comienzan a separarse, excepto en los quiasmas. Al ocurrir esto, termina el paquiteno y comienza el diploteno.
La asociación entre el recombinosoma y los ejes del complejo sinaptonémico persiste a lo largo de toda la sinapsis. Particularmente en los entrecruzamientos recombinogénicos hasta el final del paquiteno, o un poco más allá.
- Alberts, B., Johnson, A. D., Lewis, J., Morgan, D., Raff, M., Roberts, K., Walter, P. (2014) Molecular Biology of the Cell (6th Edition). W. W. Norton & Company, New York, NY, USA.
- de Massy, B. (2013) Initiation of meiotic recombination: how and where? Conservation and specificities among eukaryotes. Annual Reviews of Genetics 47, doi: 10.1146/annurev-genet-110711-155423
- Goodenough, U. W. (1984) Genetics. W. B. Saunders Co. Ltd, Philadelphia, PA, USA.
- Griffiths, A. J. F., Wessler, R., Carroll, S. B., Doebley, J. (2015). An Introduction to Genetic Analysis (11th ed.). New York: W. H. Freeman, New York, NY, USA.
- Zickler, D., Kleckner, N. (2015) Recombination, pairing, and synapsis of homologs during meiosis. Cold Spring Harbor Perspectives in Biology, doi: 10.1101/cshperspect.a016626