Canales iónicos: estructura, funciones, tipos
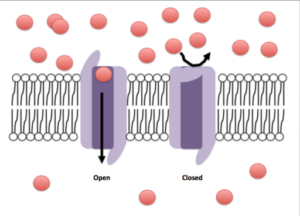
Los canales iónicos son unas estructuras membranales huecas que forman conductos o poros que atraviesan el espesor de la membrana y comunican el exterior de la célula con su citosol y viceversa; algunos pueden disponer de un sistema de compuerta que regula su apertura.
Estos canales están llenos de agua y controlan el paso de iones específicos de un lado al otro de la membrana. Están formados por unas proteínas propias de las membranas celulares que forman estructuras cilíndricas en forma de tubo que las atraviesan a lo ancho.
Los mecanismos de transporte a través de estas membranas se pueden clasificar ampliamente en transportes pasivos o activos. Los pasivos son aquellos que permiten el paso de sustancias a favor de sus gradientes de concentración, entretanto los activos requieren del consumo de energía, ya que desplazan sustancias en contra de sus gradientes de concentración.
Los canales iónicos constituyen un mecanismo de transporte pasivo que puede clasificarse de acuerdo con su especificidad, es decir, de acuerdo con el tipo de ion que dejan pasar, o en función de la forma en la que se abren o se cierran.

La función principal de estos sistemas de transporte membranales es la de permitir el paso regulado de sustancias hacia dentro o fuera de las células y mantener así las concentraciones intracelulares de iones y otras sustancias.
La presencia de las membranas celulares y los canales iónicos en fundamental para el mantenimiento de las diferencias de concentración entre los medios intracelular y extracelular, que tiene relevancia desde muchos puntos de vista.
Los canales iónicos, especialmente aquellos ligando dependientes, son muy importantes en la farmacología y la medicina, ya que muchos fármacos pueden mimetizar las funciones de los ligandos naturales y unirse así al canal, abriéndolo o cerrándolo, según sea el caso.
Otros fármacos son capaces de bloquear el sitio de unión y así impedir la acción del ligando natural.
Índice del artículo
- 1 Estructura
- 2 Funciones
- 3 Generalidades del transporte a través de la membrana
- 4 Tipos de canales iónicos
- 5 Referencias
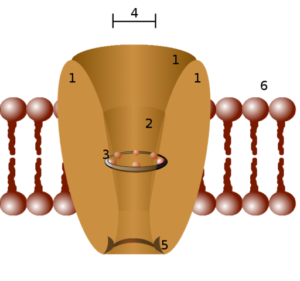
La estructura de los canales iónicos está formada por proteínas transmembranales específicas que tienen una forma tubular y dejan un poro u orificio que permite la comunicación entre el interior y el exterior de la célula o entre compartimientos intracelulares (orgánulos).
Cada canal iónico implica una proteína estructural específica de la membrana y se han descrito más de 100 genes que codifican canales iónicos específicos.
Para el canal de sodio, por ejemplo, se han descrito 10 genes llamados SCN que codifican diferentes proteínas distribuidas en distintos tejidos con funciones y estructuras particulares.
Así mismo, se ha descrito una cantidad considerable de genes que codifican distintas proteínas que conforman canales de potasio que pertenecen a diferentes familias y poseen diferentes mecanismos de activación, apertura e inactivación.
Típicamente, un canal iónico funcional asociado a una membrana está compuesto por el ensamblaje de 4 a 6 subunidades polipeptídicas similares (homo oligómeros) o diferentes (hetero oligómeros) que forman un poro central entre ellas.
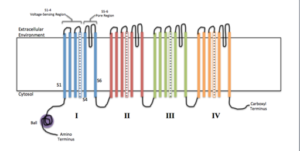
Cada subunidad varía de acuerdo con las características y propiedades del canal, ya que muchos son específicos para iones determinados y poseen mecanismos diversos de apertura y cierre.
Algunos canales están conformados por una sola cadena polipeptídica organizada en motivos repetidos que van atravesando varias veces el espesor de la membrana y funcionan como el equivalente a una subunidad proteica.
Además de estas subunidades, que se conocen en la literatura como subunidades α, algunos canales iónicos también poseen una o más subunidades auxiliares (ß o γ) que regulan la apertura y el cierre de los mismos.
La especificidad de cada canal está relacionada con el diámetro del poro formado por las proteínas transmembranales y las cadenas laterales (─R) de los aminoácidos que las conforman.
De esta manera, existen canales que solo dejan pasar iones de sodio, de potasio, de calcio y así sucesivamente, pues las cadenas laterales funcionan como “tamiz”.
Otra característica importante de muchos canales son las compuertas. Los canales con estas propiedades pueden abrirse o cerrarse frente a cambios locales que ocurren en el microambiente membranal que rodea al canal.
Dependiendo del tipo de canal, estos cambios pueden ser mecánicos, térmicos (cambios de temperatura), eléctricos (cambios de voltaje) o químicos (unión de un ligando).
No obstante, en los llamados canales iónicos pasivos, que son aquellos que permanecen abiertos y permiten el paso específico de iones determinados, estas estructuras no tienen compuertas ni son sensibles a ligandos u otro tipo de estímulos.
En otros canales iónicos, que son sensibles a la presencia o unión de ligandos, hay un sitio de unión para ligando bien sea en el lado extracelular o hacia el citosol celular y en estos casos los poros o canales poseen una compuerta que puede abrirse o cerrarse de acuerdo con el estado de su ligando.
En el caso de tener un sitio para ligando en la porción intracelular, estos canales suelen tener segundos mensajeros como ligandos. Un ejemplo de los canales iónicos que se abren o se cierran por mecanismos de segundos mensajeros es el de los receptores olfativos:
Las moléculas odoríferas se unen a sus receptores en el lado extracelular. Estos receptores, a su vez, están unidos a una proteína G que se activa que, a su vez, activa a la proteína adenilciclasa que forma AMPc, que es un segundo mensajero.
El AMPc se une a un sitio de unión intracelular de unos canales de calcio, lo que resulta en su apertura y la entrada de calcio hacia la célula.
Como si de un efecto dominó se tratase, el calcio se une a un sitio para ligando de otro canal de cloro, lo que genera su apertura y la salida de este ion, ocasionando la despolarización de la célula olfatoria.
Es importante destacar que los cambios generados por los ligandos o los estímulos que afectan a los canales iónicos corresponden a cambios conformacionales de las proteínas que constituyen la estructura del canal.
En otras palabras, los cambios conformacionales que pueden mover una compuerta y cerrar o abrir un canal, no son más que el acercamiento o el distanciamiento de las subunidades proteicas que lo componen.
Algunos canales, sobre todo los canales voltaje-dependientes, pueden entrar en un estado refractario durante el cual el mismo cambio de voltaje que los activó ahora ya no los activa.
Por ejemplo, en los canales de calcio voltaje-dependientes, el cambio de voltaje abre el canal y el calcio entra y, una vez dentro de la célula, el mismo ion se une a un sitio de unión del canal de calcio y este se cierra.
Otra forma de inactivación reversible del canal de calcio que explica su refractariedad después de la activación es la desfosforilación del canal debido al aumento de la concentración interna del calcio.
Es decir, un canal de calcio puede inactivarse irreversiblemente debido a la presencia de concentraciones patológicamente altas del ion, que median el reclutamiento de enzimas desdobladoras de otras proteínas activadas por calcio.
Los canales regulados por ligando pueden entrar en estado refractario cuando se exponen de forma prolongada a su ligando, recibiendo este mecanismo el nombre de desensibilización.
Los fármacos, los venenos y las toxinas pueden incidir sobre la regulación de los canales iónicos, cerrándolos o manteniéndolos abiertos o, en algunos casos, ocupando el sitio del ligando e interfiriendo así con su función.
Los canales iónicos tienen multiplicidad de funciones, directas o indirectas.
– Son los responsables de regular el flujo de iones a través de las membranas plasmáticas y organelares de todas las células.
– Permiten la existencia de un control sobre las concentraciones intracelulares de los diferentes iones.
– En las neuronas y en las células musculares, los canales iónicos controlan las variaciones del potencial de membrana que ocurren durante los potenciales de acción y durante los potenciales post sinápticos de las células efectoras.
– Los canales de calcio que generan flujos netos de calcio hacia el espacio intracelular son responsables de la activación de numerosas enzimas y proteínas que participan en muchos procesos metabólicos.
– Así mismo el incremento de calcio debido a un aumento en su transporte inicia el mecanismo de liberación de los neurotransmisores al espacio sináptico de las neuronas.
– Por lo tanto, la función de los canales iónicos también se relaciona con los mecanismos de comunicación celular.
Como se enunció anteriormente, los mecanismos de transporte membranales pueden ser activos o pasivos según consuman o no energía de la célula donde se encuentran. Los mecanismos pasivos se clasifican en difusión simple y difusión facilitada.
La difusión simple permite el paso a través de la estructura de fosfolípidos de la membrana de moléculas liposolubles de pequeño tamaño, con características apolares y sin carga.
Así, por ejemplo, los gases como el oxígeno (O2) y el dióxido de carbono (CO2), el etanol y la urea, por citar algunos, pasan a favor de su gradiente de concentración.
La difusión facilitada es aquella que es facilitada por proteínas y de este mecanismo de transporte pasivo existen dos tipos: los canales iónicos y las proteínas de transporte o proteínas transportadoras.
Los canales iónicos son el mecanismo más empleado por las células para el transporte de iones que no pueden pasar por difusión simple, bien sea porque tienen carga eléctrica y los fosfolípidos de la membrana los repelen, por su tamaño y polaridad o cualquier otra característica.
La difusión facilitada por proteínas transportadoras es empleada para el transporte de sustancias más grandes con o sin carga, como por ejemplo la glucosa y otros azúcares.
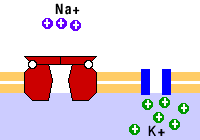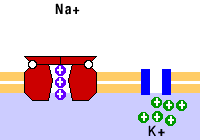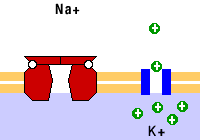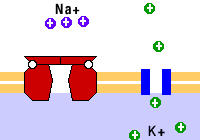
El transporte membranal activo es aquel que ocurre en contra del gradiente de concentración del soluto que se transporta y requiere del consumo de energía en forma de ATP. Entre los transportadores de este tipo están las bombas y el transporte vesicular.
Como ejemplo de las bombas está la de sodio/potasio, que saca tres sodios e introduce dos potasios. También están las bombas de calcio.
Como ejemplo del transporte vesicular están la endocitosis, la exocitosis, la pinocitosis y la fagocitosis; todos estos mecanismos de transporte activo.
A partir de este punto se hará referencia a los canales iónicos que permiten el paso de iones a través de una membrana a favor de sus gradientes de concentración, es decir, que son canales de transporte pasivo.
Generalmente, cada uno de estos canales es específico para un solo ion, con la salvedad de algunos pocos canales que permiten el transporte de pares de iones.
Una forma de clasificar los canales iónicos es agrupándolos de acuerdo con el mecanismo responsable de su apertura. Así, se han descrito los canales pasivos, los canales regulados por el voltaje (voltaje-dependientes), los canales regulados por ligando y los canales regulados por estímulos mecánicos.
– Canales pasivos: son canales que se encuentran permanentemente abiertos y no responden a ningún tipo de estímulo; estos son específicos para determinados iones.
– Canales voltaje-dependientes: estos pueden abrirse o cerrarse (dependiendo del canal) frente a cambios en el voltaje de la membrana. Son muy importantes para la señalización celular, sobretodo en el sistema nervioso central de los mamíferos.
– Canales ligando-dependientes: llamados también canales con puerta de ligando o regulados por ligando, están ampliamente distribuidos en las diferentes células corporales de los humanos, pero en el sistema nervioso constituyen aquellos canales iónicos activados por neurotransmisores y son esenciales para la transmisión sináptica y la señalización intercelular.
Ejemplo de canales iónicos ligando-dependientes activados por neurotransmisores son los canales de sodio/potasio activados por glutamato.
La activación de los receptores colinérgicos, en este caso la unión de la acetilcolina a la membrana postsináptica (ligando del canal), abre canales de sodio ligando-dependientes y permite la entrada de este ion siguiendo su gradiente de concentración.
– Canales regulados por estímulos mecánicos: son canales que se pueden activar por distensión o presión. Estas fuerzas mecánicas se transmiten al canal a través del citoesqueleto y el canal se abre.
- Bear, M. F., Connors, B. W., & Paradiso, M. A. (Eds.). (2007). Neuroscience (Vol. 2). Lippincott Williams & Wilkins.
- Department of Biochemistry and Molecular Biophysics Thomas Jessell, Siegelbaum, S., & Hudspeth, A. J. (2000). Principles of neural science (Vol. 4, pp. 1227-1246). E. R. Kandel, J. H. Schwartz, & T. M. Jessell (Eds.). New York: McGraw-hill.
- Lewis, C. A., & Stevens, C. F. (1983). Acetylcholine receptor channel ionic selectivity: ions experience an aqueous environment. Proceedings of the National Academy of Sciences, 80(19), 6110-6113.
- Nelson, D. L., Lehninger, A. L., & Cox, M. M. (2008). Lehninger principles of biochemistry. Macmillan.
- Rawn, J. D. (1998). Biochemistry. Burlington, Massachusetts: Neil Patterson Publishers.
- Viana, F., de la Peña, E., & Belmonte, C. (2002). Specificity of cold thermo transduction is determined by differential ionic channel expression. Nature neuroscience, 5(3), 254.