Cromatina: tipos, características, estructura, funciones
La cromatina es el complejo formado por ADN y proteínas, único en organismos eucariotas. En términos de proporción contiene casi el doble de proteínas que de material genético. Las proteínas más importantes de este complejo son las histonas – proteínas pequeñas con carga positiva que se unen al ADN mediante interacciones electrostáticas. Adicionalmente, la cromatina posee más de mil proteínas diferentes a las histonas.
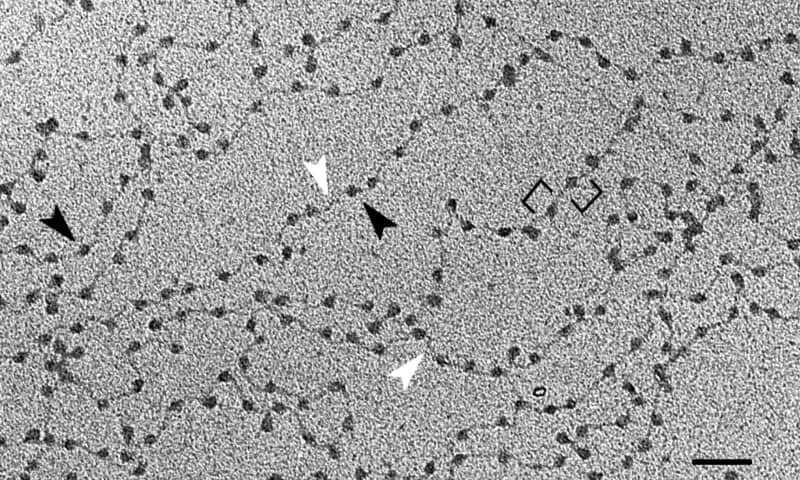
La unidad fundamental de la cromatina es el nucleosoma, el cual consiste en la unión de histonas y ADN. Esta ordenación recuerda a las cuentas de un collar. Tras pasar por todos los niveles de organización superiores del ADN, llegaremos a los cromosomas.
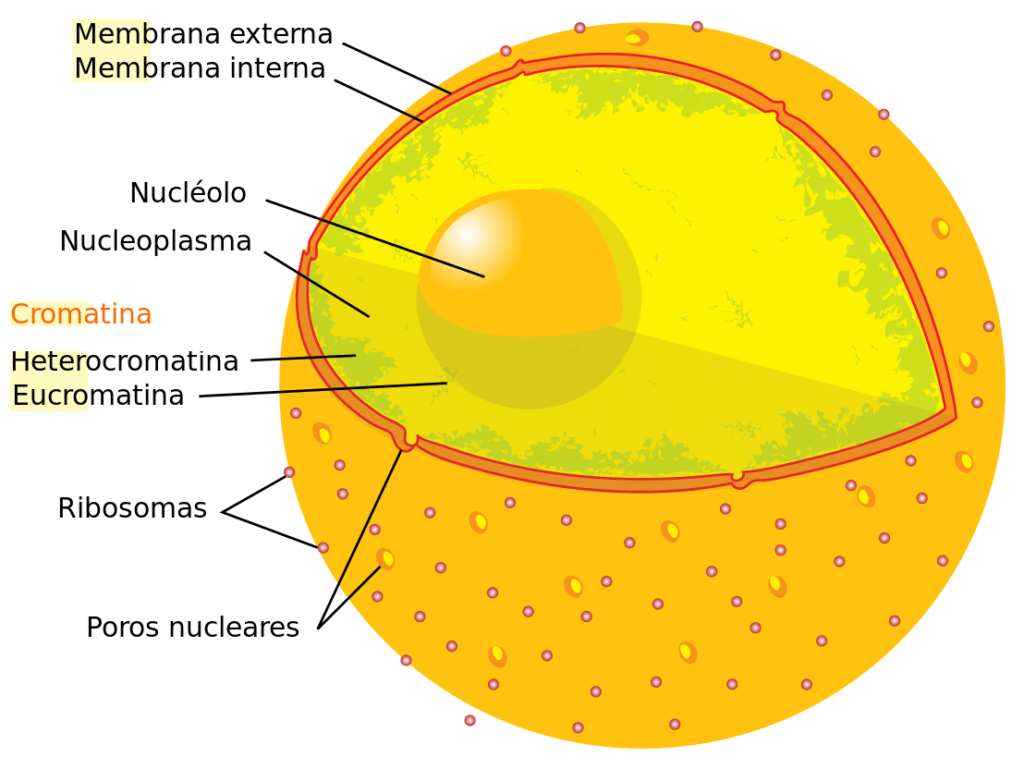
La estructura de la cromatina está íntimamente relacionada con el control de la expresión génica. Existen dos tipos principales: la eucromatina y la heterocromatina.
La eucromatina se caracteriza por tener un bajo grado de compactación, lo que se traduce en altos niveles de transcripción. En contraste, la heterocromatina es transcripcionalmente inactiva, debido al grado de compactación tan elevado que presenta.
Estructuralmente existen ciertas marcas epigenéticas de las histonas propias de ambos tipos de cromatinas. Mientras que la eucromatina se asocia con la acetilación, la heterocromatina se asocia con la disminución en grupos químicos.
Existen algunas regiones de heterocromatina con funciones estructurales, como los telómeros y los centrómeros.
Índice del artículo
El estudio del material genético y su organización estructural empieza en 1831 cuando el investigador Robert Brown describe el núcleo. Una de las preguntas inmediatas a este descubrimiento fue indagar acerca de la naturaleza biológica y química de dicha estructura.
Estas preguntas empezaron a ser dilucidadas entre 1870 y 1900, con los experimentos de Friedrich Miescher, quien introduce la palabra nucleina. Sin embargo, Walther Flemming modifica el término y usa cromatina para referirse a la sustancia nuclear.
Con el paso del tiempo se empieza a tener un conocimiento más profundo del material genético y sus propiedades. No es hasta 1908 cuando el investigador oriundo de Italia Pasquale Baccarini nota que la cromatina no es homogénea y logra visualizar pequeños cuerpos en el interior del núcleo.
Los tipos de cromatinas – eucromatina y heterocromatina – fueron inicialmente propuestos por Emil Heitz en 1928. Para establecer dicha clasificación, Heitz se basó en el uso de tinciones.
En 1974 el biólogo Roger Kornberg propuso un modelo para la organización del material genética en estructuras conocidas como nucleosomas, hipótesis confirmada empíricamente por los experimentos de Markus Noll.
La cromatina es una nucleoproteína formada por la unión del material genético – el ADN – con un conjunto heterogéneo de proteínas. Esta asociación es altamente dinámica y adquiere un conformación tridimensional compleja que le permite llevar a cabo sus funciones regulatorias y estructurales.
Una de las proteínas más importante de la cromatina son las histonas, las cuales se encuentran casi en igual proporción que el ADN.
Las histonas son proteínas básicas, notablemente conservadas a lo largo de la historia evolutiva de los seres orgánicos – es decir, nuestras histonas no varían mucho comparadas con las de otros mamíferos, incluso otro animal más lejano filogenéticamente.
La carga de las histonas es positiva, por lo que pueden interactuar por medio de fuerzas electrostáticas con la carga negativa del esqueleto de fosfatos presente en el ADN. Existen cinco tipos de histonas, a saber: H1, H2A, H2B, H3, and H4.
También existen una serie de proteínas de naturaleza diferente a las histonas que participan en la compactación del ADN.
La unidad básica de la cromatina son los nucleosomas – estructuras repetitivas formadas de ADN e histonas, conformación que encontramos a lo largo del material genético.
La doble hélice de ADN se enrolla en un complejo de ocho histonas conocido como octámero de histonas. La molécula se enrolla en dos vueltas aproximadamente, seguidos por una región corta (entre 20 y 60 pares de bases) que separa a los nucleosomas unos de otros.
Para entender esta organización, debemos tomar en cuenta que la molécula de ADN es extremadamente larga (unos 2 metros) y debe enrollarse de manera ordenada para establecerse en el núcleo (cuyo diámetro es de 3 a 10 µm). Además, debe quedar disponible para su replicación y transcripción.
Este objetivo se logra con distintos niveles de compactación del ADN, siendo el primero de ellos los mencionados nucleosomas. Estos se asemejan a las cuentas de un collar de perlas. Aproximadamente 150 pares de bases de ADN se enrollan en casa “cuenta”.
En las bacterias no existen historias verdaderas. En contraste, existen una serie de proteínas que recuerdan a las histonas y se presume que estas contribuyen en el empaquetamiento del ADN bacteriano.
La organización de lo cromatina no se restringe a nivel de los nucleosomas. Esta asociación de proteínas y ADN se agrupan en una estructura más gruesa de unos 30 nm – debido a este grosor se le denomina al nivel “fibra de 30 nm”.
La cromatina organizada en el grosor de 30 nm se organiza a su vez en forma de bucles que se extienden en una especie de andamio de naturaleza proteica (no histonas).
Este es el modelo que se maneja actualmente, aunque se puede esperar la existencia de mecanismos de compactación más complejos. La organización final consiste en el cromosoma.
La compactación y organización del material genético es vital para múltiples funciones biológicas. Distintas condiciones médicas han sido asociadas con errores en la estructura de la cromatina, entre ellos alfa talasemia ligada al cromosoma X, síndrome de Rubinstein-Taybi, síndrome de Coffin-Lowry, síndrome de Rett, entre otros.
Existen dos tipos de cromatina en la célula, revelada por la aplicación de tinciones: la eucromatina (cromatina “verdadera”) y la heterocromatina. En el primer caso la tinción se observa débilmente, mientras que en el segundo la tinción es intensa.
Esta organización estructural del ADN es única de los organismos eucariotas y es crucial para el comportamiento de los cromosomas y la regulación de la expresión génica.
Si evaluamos las proporciones de ambos tipos de cromatinas en una célula que se encuentra en interfase encontramos que aproximadamente el 90% de la cromatina es eucromatina y el 10% restante corresponde a heterocromatina. A continuación describiremos con detalle cada tipo:
Características
La diferencia principal que existe entre ambos tipos de cromatina está relacionada con el grado de compactación o “empaquetamiento” de la molécula durante etapas puntuales en la división celular.
Aunque en la interfase el material genético pareciera estar disperso al azar, no es de esta manera.
Existe una organización significativa en esta etapa, donde se puede apreciar una partición diferencial del material cromosómico en el interior del núcleo.
El ADN de los cromosomas no se entrelaza con la hebra de ADN de otros cromosomas y permanece es regiones específicas denominadas territorios cromosomales. Esta organización pareciera contribuir en la expresión génica.
La heterocromatina se encuentra condensada fuertemente, quedando poco accesible para la maquinaria de transcripción – por lo que no se transcriben. Además, es pobre en términos de la cantidad de genes que presenta.
Tipos de heterocromatina
Ciertas regiones de heterocromatina son persistentes a lo largo de los linajes celulares – es decir, siempre se comportará como heterocromatina. Este tipo de heterocromatina se conoce como constitutiva. Ejemplo de ello son las regiones condensadas de los cromosomas denominados centrómeros y telómeros.
En contraste, existen porciones de heterocromatina que pueden variar el nivel de compactación en respuesta a cambios en los patrones de desarrollo o a variables ambientales.
Gracias a los nuevos estudios esta visión está siendo reformulada y ahora existe evidencia de que la heterocromatina constitutiva también es dinámica y capaz de responder a estímulos.
Estructura
Uno de los factores que determina la estructura de la cromatina son las modificaciones químicas de las histonas. En el caso de la cromatina que es transcripcionalmente inactiva exhiben unas histonas hipoacetiladas.
La disminución en la cantidad de grupos acetilo se asocia con el silenciamiento de genes, ya que no enmascara la carga positiva de las lisinas, permitiendo una fuerte interacción electrostática entre el ADN y las histonas.
Otra marca epigenética es la metilación. Sin embargo, como la añadidura de un grupo metilo no modifica la carga de la proteína, su consecuencia (activar o desactivar genes) no es tan obvia y dependerá de la región de la histona donde se encuentre la marca.
Empíricamente se ha encontrado que la metilación de H3K4me3 y H3K36me3 se asocia con activación génica y las de H3K9me3 y H3K27me3.
Funciones
En el ejemplo de la heterocromatina constitutiva mencionamos al centrómero. Esta región cromosómica tiene un papel estructural y contribuye al movimiento de los cromosomas durante los eventos de división celular tanto mitóticos como meióticos.
Características
Contrario a la heterocromatina, la eucromatina es una molécula menos compacta, por lo que la maquinaria de transcripción tiene fácil acceso (específicamente a la enzima ARN polimerasa) y puede expresarse por vías genéticas activamente.
Estructura
La estructura de un nucleosoma de una cromatina activa desde el punto de vista de la transcripción se caracteriza por histonas acetiladas. La presencia de lisinas monometiladas también se asocian con activación de genes.
La adición de un grupo acetilo a estos residuos de lisina de las histonas neutraliza la carga positiva de dicho aminoácido. La consecuencia inmediata de este cambio es la reducción de las interacciones electrostáticas entre la histona y el ADN, generando una cromatina más laxa.
Esta modificación estructural permite las interacciones del material genético con la maquinaria transcripcional, la cual se caracteriza por ser particularmente abultada.
Funciones
La eucromatina engloba a todos los genes que se encuentra activos y logran ser alcanzados por la maquinaria enzimática relacionada a la transcripción. Por ello, las funciones son tan amplias como las funciones de los genes implicados.
- Grewal, S. I., & Moazed, D. (2003). Heterochromatin and epigenetic control of gene expression. science, 301(5634), 798-802.
- Jost, K. L., Bertulat, B., & Cardoso, M. C. (2012). Heterochromatin and gene positioning: inside, outside, any side?. Chromosoma, 121(6), 555–563.
- Lewin, B. (2008). Genes IX. Jones and Bartlett Publishers.
- Tollefsbol, T. O. (2011). Handbook of Epigenetics. Academic Press.
- Wang, J., Jia, S. T., & Jia, S. (2016). New Insights into the Regulation of Heterochromatin. Trends in genetics : TIG, 32(5), 284–294.
- Zhang, P., Torres, K., Liu, X., Liu, C. G., & Pollock, R. E. (2016). An Overview of Chromatin-Regulating Proteins in Cells. Current protein & peptide science, 17(5), 401–410.